State of the Physical, Biological and Selected Fishery Resources of Pacific Canadian Marine Ecosystems in 2016
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-
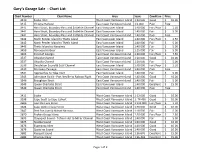
Gary's Charts
Gary’s Garage Sale - Chart List Chart Number Chart Name Area Scale Condition Price 3410 Sooke Inlet West Coast Vancouver Island 1:20 000 Good $ 10.00 3415 Victoria Harbour East Coast Vancouver Island 1:6 000 Poor Free 3441 Haro Strait, Boundary Pass and Sattelite Channel East Vancouver Island 1:40 000 Fair/Poor $ 2.50 3441 Haro Strait, Boundary Pass and Sattelite Channel East Vancouver Island 1:40 000 Fair $ 5.00 3441 Haro Strait, Boundary Pass and Sattelite Channel East Coast Vancouver Island 1:40 000 Poor Free 3442 North Pender Island to Thetis Island East Vancouver Island 1:40 000 Fair/Poor $ 2.50 3442 North Pender Island to Thetis Island East Vancouver Island 1:40 000 Fair $ 5.00 3443 Thetis Island to Nanaimo East Vancouver Island 1:40 000 Fair $ 5.00 3459 Nanoose Harbour East Vancouver Island 1:15 000 Fair $ 5.00 3463 Strait of Georgia East Coast Vancouver Island 1:40 000 Fair/Poor $ 7.50 3537 Okisollo Channel East Coast Vancouver Island 1:20 000 Good $ 10.00 3537 Okisollo Channel East Coast Vancouver Island 1:20 000 Fair $ 5.00 3538 Desolation Sound & Sutil Channel East Vancouver Island 1:40 000 Fair/Poor $ 2.50 3539 Discovery Passage East Coast Vancouver Island 1:40 000 Poor Free 3541 Approaches to Toba Inlet East Vancouver Island 1:40 000 Fair $ 5.00 3545 Johnstone Strait - Port Neville to Robson Bight East Coast Vancouver Island 1:40 000 Good $ 10.00 3546 Broughton Strait East Coast Vancouver Island 1:40 000 Fair $ 5.00 3549 Queen Charlotte Strait East Vancouver Island 1:40 000 Excellent $ 15.00 3549 Queen Charlotte Strait East -

A Salmon Monitoring & Stewardship Framework for British Columbia's Central Coast
A Salmon Monitoring & Stewardship Framework for British Columbia’s Central Coast REPORT · 2021 citation Atlas, W. I., K. Connors, L. Honka, J. Moody, C. N. Service, V. Brown, M .Reid, J. Slade, K. McGivney, R. Nelson, S. Hutchings, L. Greba, I. Douglas, R. Chapple, C. Whitney, H. Hammer, C. Willis, and S. Davies. (2021). A Salmon Monitoring & Stewardship Framework for British Columbia’s Central Coast. Vancouver, BC, Canada: Pacific Salmon Foundation. authors Will Atlas, Katrina Connors, Jason Slade Rich Chapple, Charlotte Whitney Leah Honka Wuikinuxv Fisheries Program Central Coast Indigenous Resource Alliance Salmon Watersheds Program, Wuikinuxv Village, BC Campbell River, BC Pacific Salmon Foundation Vancouver, BC Kate McGivney Haakon Hammer, Chris Willis North Coast Stock Assessment, Snootli Hatchery, Jason Moody Fisheries and Oceans Canada Fisheries and Oceans Canada Nuxalk Fisheries Program Bella Coola, BC Bella Coola, BC Bella Coola, BC Stan Hutchings, Ralph Nelson Shaun Davies Vernon Brown, Larry Greba, Salmon Charter Patrol Services, North Coast Stock Assessment, Christina Service Fisheries and Oceans Canada Fisheries and Oceans Canada Kitasoo / Xai’xais Stewardship Authority BC Prince Rupert, BC Klemtu, BC Ian Douglas Mike Reid Salmonid Enhancement Program, Heiltsuk Integrated Resource Fisheries and Oceans Canada Management Department Bella Coola, BC Bella Bella, BC published by Pacific Salmon Foundation 300 – 1682 West 7th Avenue Vancouver, BC, V6J 4S6, Canada www.salmonwatersheds.ca A Salmon Monitoring & Stewardship Framework for British Columbia’s Central Coast REPORT 2021 Acknowledgements We thank everyone who has been a part of this collaborative Front cover photograph effort to develop a salmon monitoring and stewardship and photograph on pages 4–5 framework for the Central Coast of British Columbia. -

Uvic Thesis Template
Coastal aquaculture in British Columbia: Perspectives on finfish, shellfish, seaweed and Integrated Multi-Trophic Aquaculture (IMTA) from three First Nation communities by Kathryn Tebbutt B.A., University of British Columbia, 2009 A Thesis Submitted in Partial Fulfillment of the Requirements for the Degree of MASTER OF ARTS in the Department of Geography Kathryn Tebbutt, 2014 University of Victoria All rights reserved. This thesis may not be reproduced in whole or in part, by photocopy or other means, without the permission of the author. ii Supervisory Committee Coastal aquaculture in British Columbia: Perspectives on finfish, shellfish, seaweed and Integrated Multi-Trophic Aquaculture (IMTA) from three First Nation communities by Kathryn Tebbutt B.A., University of British Columbia, 2009 Supervisory Committee Dr. Mark Flaherty, (Department of Geography) Supervisor Dr. Denise Cloutier, (Department of Geography) Departmental Member Dr. Stephen Cross, (Department of Geography) Departmental Member iii Abstract Supervisory Committee Dr. Mark Flaherty, (Department of Geography) Supervisor Dr. Denise Cloutier, (Department of Geography) Departmental Member Dr. Stephen Cross, (Department of Geography) Departmental Member Most aquaculture tenures in British Columbia (BC) are located in coastal First Nation traditional territories, making the aquaculture industry very important to First Nation communities. Marine aquaculture, in particular salmon farming, has been labeled one of the most controversial industries in BC and various groups with differing opinions have created a wide-spread media debate known as the “aquaculture controversy”. Industry, government, and (E)NGO’s are often the most visible players; First Nations, especially those without aquaculture operations directly in their territories, are often excluded or underrepresented in the conversation. -

The Toxicological Effects of the Mount Polley Tailings Impoundment Breach on Freshwater Amphipods
THE TOXICOLOGICAL EFFECTS OF THE MOUNT POLLEY TAILINGS IMPOUNDMENT BREACH ON FRESHWATER AMPHIPODS RAEGAN D. PLOMP Bachelor of Science, University of Lethbridge, 2017 A thesis submitted in partial fulfilment of the requirements for the degree of MASTER OF SCIENCE in BIOLOGICAL SCIENCES Department of Biological Sciences University of Lethbridge LETHBRIDGE, ALBERTA, CANADA © Raegan Plomp, 2019 THE TOXICOLOGICAL EFFECTS OF THE MOUNT POLLEY TAILINGS IMPOUNDMENT BREACH ON FRESHWATER AMPHIPODS RAEGAN D. PLOMP Date of Defence: September 6, 2019 Dr. G. Pyle Professor Ph.D. Supervisor Dr. S. Wiseman Associate Professor Ph.D. Thesis Examination Committee Member Dr. R. Laird Associate Professor Ph.D. Thesis Examination Committee Member Dr. Laura Chasmer Assistant Professor Ph.D. Internal External Examiner Department of Geography Dr. T. Russell Associate Professor Ph.D. Chair, Thesis Examination Committee ii Abstract The bioavailability and toxicity of metals to amphipods is influenced by exposure route and co-toxic mechanisms. Metal bioavailability was studied in amphipods at sites affected by the 2014 Mount Polley Mining Corporation tailings impoundment breach. The area around the lake was also subjected to wildfire in 2017. Bioavailability was more correlated with sediment than waterborne metal concentrations with Cu correlating well with distance from the breach. Copper-sediment bioavailability to amphipods in combination with wildfire runoff was studied resulting in non-additive toxicity. Hyalella azteca exposed to Cu-enriched sediment with fire extract (FE) experienced a more-than- additive effect on survival and amphipod whole-body Cu concentration but no significant reduction in growth or acetylcholinesterase activity compared to the Cu-contaminated sediment or FE alone, respectively. -

Burrard Inlet Underwater Noise Study
Vancouver Fraser Port Authorit Burrard Inlet underwater noise study: 2020 final report ECHO Program study summary This study was undertaken for the Vancouver Fraser Port Authority-led Enhancing Cetacean Habitat and Observation (ECHO) Program and project partner Tsleil-Waututh Nation with financial support from Transport Canada to learn more about underwater noise and cetacean presence in Burrard Inlet. Building upon the 2019 monitoring project in Burrard Inlet, this project set out to monitor underwater noise and the presence of cetaceans (whales, dolphins and porpoise) in Burrard Inlet, a marine mammal habitat and a key waterway for commercial shipping, port-related activities, and passenger transportation. This document summarizes the project question and describes the methods, key findings, and conclusions. What questions was the study trying to answer? The second year of the Burrard Inlet underwater noise study sought to evaluate longer-term trends in total ambient noise and marine mammal presence, while building upon the results from 2019. Who conducted the project? SMRU Consulting North America (SMRU) was awarded the contract for the 2019 monitoring program, and was retained by Vancouver Fraser Port Authority to continue monitoring though 2020 at fewer sampling locations. What methods were used? Bottom-mounted SoundTrap hydrophone recorders were deployed in two locations in the inner and outer harbour: one at Burrard Inlet East near the Tsleil-Waututh Nation reserve lands and Burnaby petroleum terminals, and one in English Bay between anchorages 1 and 3. Acoustic data were collected over approximately one year between February 2020 and February 2021. The figure below shows the approximate locations of the hydrophone deployments. -

We Are the Wuikinuxv Nation
WE ARE THE WUIKINUXV NATION WE ARE THE WUIKINUXV NATION A collaboration with the Wuikinuxv Nation. Written and produced by Pam Brown, MOA Curator, Pacific Northwest, 2011. 1 We Are The Wuikinuxv Nation UBC Museum of Anthropology Pacific Northwest sourcebook series Copyright © Wuikinuxv Nation UBC Museum of Anthropology, 2011 University of British Columbia 6393 N.W. Marine Drive Vancouver, B.C. V6T 1Z2 www.moa.ubc.ca All Rights Reserved A collaboration with the Wuikinuxv Nation, 2011. Written and produced by Pam Brown, Curator, Pacific Northwest, Designed by Vanessa Kroeker Front cover photographs, clockwise from top left: The House of Nuakawa, Big House opening, 2006. Photo: George Johnson. Percy Walkus, Wuikinuxv Elder, traditional fisheries scientist and innovator. Photo: Ted Walkus. Hereditary Chief Jack Johnson. Photo: Harry Hawthorn fonds, Archives, UBC Museum of Anthropology. Wuikinuxv woman preparing salmon. Photo: C. MacKay, 1952, #2005.001.162, Archives, UBC Museum of Anthropology. Stringing eulachons. (Young boy at right has been identified as Norman Johnson.) Photo: C. MacKay, 1952, #2005.001.165, Archives, UBC Museum of Anthropology. Back cover photograph: Set of four Hàmac! a masks, collection of Peter Chamberlain and Lila Walkus. Photo: C. MacKay, 1952, #2005.001.166, Archives, UBC Museum of Anthropology. MOA programs are supported by visitors, volunteer associates, members, and donors; Canada Foundation for Innovation; Canada Council for the Arts; Department of Canadian Heritage Young Canada Works; BC Arts Council; Province of British Columbia; Aboriginal Career Community Employment Services Society; The Audain Foundation for the Visual Arts; Michael O’Brian Family Foundation; Vancouver Foundation; Consulat General de Vancouver; and the TD Bank Financial Group. -

Clayoquot Sound)
.. Catalogue of Salmon Streams and Spawning Escapements of Statistical Area 24 ( Clayoquot Sound) R.F Brown, M.J. Comfort, & D.E. Marshall . Fisheries &Oceans Enhancement Services Branch 1090 West Pender St. Vancouver. B. C. V6E 2P1 December 1979 Fisheries & Marine Service Data Report No. 80 Fisheries and Marine Service Data Reports These reports provide a medium for filing and archiving data compilations where little or no analysis is included. Such compilations commonly will have been prepared in support of other journal publications or reports. The subject matter of Data Reports reflects the broad interests and policies of the Fisheries and M arine Service, namely, fisheries management, technology and development, ocean sciences, and aquatic environments relevant to Canada. Numbers 1-25 in this series were issued as Fisheries and Marine Service Data Records by the Pacific Biological Station, N anaimo, B.C The series name was changed with report number 26. Data Reports are not intended for general distribution and the contents must not be referred to in other publications without prior written clearance from the Issuing establishment. The correct citation appears above the abstract of each report. Service des peches et de la mer Rapports statistiques Ces rapports servent de base a la compilation des donnees de classel11ent et d'archives pour lesquelles iI y a peu ou point d'analyse. Celte compilation aura d'ordinaire ete preparee pour appuyer d'autres publications ou rapports. Les sujets des Rapports statistiques refietent la vaste gamme des interets et politiques du Service des peches et de la mer, notamment gestion des peches, techniques et developpement, sciences oceaniques et environnements aquatiques, au Canada . -

You Are in Wolf and Cougar Country
t Keep Predators Wild & Wary ... Stay Safe! Parks Canada needs YOUR help to If you encounter a wolf or cougar: prevent people-predator conflicts and Are People at Risk? • Pick up small children. to keep our predators wild! Cougars very rarely prey on people. Children and crouching adults are more at • Gather the group together. risk of attack as they more closely resemble prey. People on their own are also • Do not run. Wolves and cougars are native to Vancouver Island, more at risk than groups of people. Wolf attacks are even rarer. • Do not crouch down. and as predators, are vital to a healthy coastal In this region, there have been three cougar attacks. In each case children were • Make and maintain eye contact. ecosystem. They may be encountered anywhere in attacked: one on the West Coast Trail in 1985, one fatal attack just outside the • Wave your arms and shout. Pacific Rim National Park Reserve. National parks are park in Clayoquot Sound in 1989 and another at Kennedy Lake in 2011. • Do all you can to appear larger and to scare the great places to view wildlife in their natural habitat. animal away. However, once animals become accustomed to In 2000, a wolf that had obtained food from previous campers, attacked a © Parks Canada / Josh McCulloch 2009 • Avoid scaring the animal into the path of other © Parks Canada / Josh McCulloch 2004 sleeping man by a campfire in Clayoquot Sound. people they are in danger of losing their wildness. Keep kids close. people. The repeated presence of humans that brings no Keep kayak hatches secure. -

Reduced Annualreport1972.Pdf
PROVINCE OF BRITISH COLUMBIA DEPARTMENT OF RECREATION AND CONSERVATION HON. ROBERT A. WILLIAMS, Minister LLOYD BROOKS, Deputy Minister REPORT OF THE Department of Recreation and Conservation containing the reports of the GENERAL ADMINISTRATION, FISH AND WILDLIFE BRANCH, PROVINCIAL PARKS BRANCH, BRITISH COLUMBIA PROVINCIAL MUSEUM, AND COMMERCIAL FISHERIES BRANCH Year Ended December 31 1972 Printed by K. M. MACDONALD, Printer to tbe Queen's Most Excellent Majesty in right of the Province of British Columbia. 1973 \ VICTORIA, B.C., February, 1973 To Colonel the Honourable JOHN R. NICHOLSON, P.C., O.B.E., Q.C., LLD., Lieutenant-Governor of the Province of British Columbia. MAY IT PLEASE YOUR HONOUR: Herewith I beg respectfully to submit the Annual Report of the Department of Recreation and Conservation for the year ended December 31, 1972. ROBERT A. WILLIAMS Minister of Recreation and Conservation 1_) VICTORIA, B.C., February, 1973 The Honourable Robert A. Williams, Minister of Recreation and Conservation. SIR: I have the honour to submit the Annual Report of the Department of Recreation and Conservation for the year ended December 31, 1972. LLOYD BROOKS Deputy Minister of Recreation and Conservation CONTENTS PAGE Introduction by the Deputy Minister of Recreation and Conservation_____________ 7 General Administration_________________________________________________ __ ___________ _____ 9 Fish and Wildlife Branch____________ ___________________ ________________________ _____________________ 13 Provincial Parks Branch________ ______________________________________________ -

Coastal Waterbird Population Trends in the Strait of Georgia 1999–2011: Results from the First 12 Years of the British Columbia Coastal Waterbird Survey
8 Coastal waterbird trends - Crewe et al. Coastal waterbird population trends in the Strait of Georgia 1999–2011: Results from the first 12 years of the British Columbia Coastal Waterbird Survey Tara Crewe1, Karen Barry2, Pete Davidson2,3, Denis Lepage1 1 Bird Studies Canada - National Headquarters, PO Box. 160, Port Rowan, Ont. N0E 1M0 2 Bird Studies Canada – British Columbia Program, 5421 Robertson Road, RR1, Delta, B.C. V4K 3N2; e-mail: [email protected] 3 Corresponding author Abstract: The British Columbia Coastal Waterbird Survey is a citizen science long-term monitoring program implemented by Bird Studies Canada to assess population trends and ecological needs of waterbirds using the province’s coastal and inshore marine habitats. Standard monthly counts from more than 200 defined sites within the Strait of Georgia were analysed using route-regression techniques to estimate population indices and assess trends in 57 waterbird species over a 12-year period spanning the non-breeding periods from 1999–2000 to 2010–11. A power analysis was also conducted to validate the rigor of the survey design. Results indicate that the survey is detecting annual changes of 3% or less for populations of 29 waterbirds of a wide variety of guilds. Thirty-three species showed stable populations or no trend, 22 species showed significantly declining trends, and just three species showed significant increasing trends. We evaluate these results in the context of other long-term monitoring initiatives in the Salish Sea, highlighting specific birds to watch from a conservation perspective. Among those that showed a declining trend were a guild of piscivores, including Western and Horned Grebes, Common, Red-throated and Pacific Loons, and Rhinoceros Auklet; several sea ducks (Black and White-winged Scoters, Long-tailed Duck, Barrow’s Goldeneye, Harlequin Duck); two shorebirds (Dunlin, Surfbird); and Great Blue Heron. -

Vital Signs Report
CLAYOQUOT SOUND BIOSPHERE REGION’S 2018 Welcome to the Clayoquot Sound Biosphere Region’s Vital Signs® 2018 Table of Contents From the Vital Signs Research Team About Vital Signs 2 “We hope the 2018 Vital Signs report ¸ Grounded in the Nuu-chah-nulth (nuucaanuł) ¸ ˇ informs and inspires dialogue and principle of hišukniš cawaak, everything is one, Vital Our Region 3 collaboration to further our collective Signs 2018 can help us to understand the complex Cycle of Poverty efforts to build healthy communities and changing systems in which we live and the necessary pathways we need to navigate in order to in Our Region: and achieve sustainable development.” Inspiring Action support sustainable ecosystems and¸ communities. One of these pathways is nuucaanułˇ language for Change 4 Tammy Dorward and Catherine Thicke revitalization. This year, we’ve worked with a regional Co-chairs, Board of Directors Environment 5-6 committee of elders¸ and language keepers to incor- Clayoquot Biosphere Trust porate nuucaanułˇ throughout the report. Climate Change Impacts 7-8 We’ve collected a range of local data to highlight pri- ority areas for community-wide action and listened People & Work 9 From our Executive Director closely to community concerns. We’ve heard that Income Inequality 10 I am pleased to present our 2018 Vital Signs report. our young people are struggling with mental health Vital Signs is a valuable tool for understanding our issues and that they lack youth programs. Families Housing 11 progress toward achieving all aspects of sustainabili- are challenged with rising housing costs and the ty—cultural, social, economic, and environmental. -

PROVINCI L Li L MUSEUM
PROVINCE OF BRITISH COLUMBIA REPORT OF THE PROVINCI_l_Li_L MUSEUM OF NATURAL HISTORY • FOR THE YEAR 1930 PRINTED BY AUTHORITY OF THE LEGISLATIVE ASSEMBLY. VICTORIA, B.C. : Printed by CHARLES F. BANFIELD, Printer to tbe King's Most Excellent Majesty. 1931. \ . To His Honour JAMES ALEXANDER MACDONALD, Administrator of the Province of British Columbia. MAY IT PLEASE YOUR HONOUR: The undersigned respectfully submits herewith the Annual Report of the Provincial Museum of Natural History for the year 1930. SAMUEL LYNESS HOWE, Pt·ovincial Secretary. Pt·ovincial Secretary's Office, Victoria, B.O., March 26th, 1931. PROVINCIAl. MUSEUM OF NATURAl. HISTORY, VICTORIA, B.C., March 26th, 1931. The Ho1Wm·able S. L. Ho11ie, ProvinciaZ Secreta11}, Victo1·ia, B.a. Sm,-I have the honour, as Director of the Provincial Museum of Natural History, to lay before you the Report for the year ended December 31st, 1930, covering the activities of the Museum. I have the honour to be, Sir, Your obedient servant, FRANCIS KERMODE, Director. TABLE OF CONTENTS . PAGE. Staff of the Museum ............................. ------------ --- ------------------------- ----------------------------------------------------- -------------- 6 Object.. .......... ------------------------------------------------ ----------------------------------------- -- ---------- -- ------------------------ ----- ------------------- 7 Admission .... ------------------------------------------------------ ------------------ --------------------------------------------------------------------------------