Acid Citrus Fruit Cultivar Improvement Via Interploid Hybridization
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

New Acid Citrus Selections for Florida
New acid citrus selections for Florida Lemon and lime-like selections with niche market potential are being developed with biotechnology at the University of Florida. By Jude Grosser, Zenaida Viloria and Manjul Dutt re you ready for a purple margarita? Would you like a fragrant, juicy lime is a naturally occurring citrus lemon for your iced tea with no seeds to clog your straw or dishwasher triploid, which is why it is seedless. drain? How about some seedless lime trees that are cold-hardy enough The new seedless watermelons in the Afor Central Florida? These and many more interesting acid-citrus marketplace are also triploids. selections are now on the horizon, including some with good ornamental potential. Due to the fact that new and This article will focus on progress in using emerging biotechnologies to develop improved citrus cultivars must be interesting new citrus cultivars in the lemon and lime group. Cultivars include seedless to compete in the national some that will not have regulatory constraints, and also a genetically modified and international marketplace, the organism (GMO)-derived purple Key lime as a teaser for the future. University of Florida’s Citrus Research and Education Center (UF/CREC) LEARNING FROM they are triploids. People and most citrus improvement team (working THE BANANA citrus trees are diploid, meaning with Fred Gmitter) has formulated Have you ever wondered why you there are two sets of chromosomes in several ways to create triploids as a key never find seeds in your bananas? Did each cell. Triploid bananas have three method of developing seedless citrus you know that there are wild-type sets of chromosomes per cell. -

Crop Profile for Citrus (Minor) Lime, Pummelo, and Kumquat in Florida
Crop Profile for Citrus (Minor) Lime, Pummelo, and Kumquat in Florida Prepared: November, 2001 General Production Information ● Lime in this crop profile refers to the 'Tahiti' lime (Citrus latifolia). Pest management practices of other members of the Rutaceae such as Key lime (Citrus aurantifolia), pummelo (Citrus grandis), and kumquat (Fortunella sp.) may also be described, since survey questionnaires grouped these four tropical fruits together. All of these species of citrus are grown for the fresh market. ● The 1997-1998 average yield of limes in Florida was 14,344 pounds per acre. At a price of $5.02 per box, the Florida crop (440,000 boxes) was worth approximately $2.21 million (1). ● In 2000-2001, 1,171 acres in Florida produced 22 million pounds of 'Tahiti' lime. This is a 58 percent decrease from the previous year, when 53 million pounds were recorded (2,3). ● As of the end of 2000, there has been a 63 percent reduction in lime acreage since 1996 (bearing acreage of 3,168 acres). Except for 20 acres, all of the lime production is located in Miami-Dade County. Consequently, ninety-eight percent of 'Tahiti' lime production occurs in this county (3). ● Florida was ranked first in the United States in lime production. No other states were listed as leading lime-producers (1). ● Reductions in lime acreage have occurred due to Hurricane Andrew and citrus canker. Bearing acreage before Andrew was approximately 6,000 acres, which dropped drastically to 1,668 acres after this event (4). An estimated 290,000 lime trees (2,800 acres) have been lost to citrus canker in the 2000-2001 period (2). -

About Limes by George Geary CCP
All About Limes By George Geary CCP Lime History In the eighteenth century, Scottish naval surgeon Sir James Lind learned by his observation of long-haul sailors that citrus fruits conquered the dreaded scurvy (lack of Vitamin C) which had divested the ranks of the British navy more than any enemy. Between 1795 and 1815, some 1.6 million gallons of lime Juice drastically reduced the mortality rate of seamen. Along with their daily ration of rum, British sailors were required to consume a daily ration of lime Juice; hence British seamen became known as limeys. Since Britain was often at war with Mediterranean countries that exported lemons, limes imported cheaply from the English colony of Jamaica were substituted as the citrus of choice. Key Lime (also known as Mexican Lime and West Indies Lime) Cultivated for thousands of years in the Indo-Malayan region, this variety has long been treasured for its fruit and decorative foliage. The Key lime made its way to North Africa and the Near East via Arabian traders, and then carried on to Palestine and Mediterranean Europe by the Crusaders. Columbus is credited with bringing the Key lime to Haiti, where Spanish settlers to Florida carried it on. It flourished in South Florida, particularly the Florida Keys, hence the current common name of Key lime. Due to hurricane-depleted soils, locals switched from pineapple commercial crops to limes in 1906, and business boomed until a hurricane once again reared and wiped out the lime groves, never to be restored. Today, most Key limes come from Mexico. -

Key Lime Pie Pollination ______
Key Lime Pie pollination ______ Headstash x Key Lime Pie Type: Sativa Finish time indoor / outdoor: 9-10 / October 5th - October 18th Yield: Medium Terp profile: Gas This hybrid yields very well in both flower and resin production. With the Key Lime Pie male you have a solid work horse combined with the award winning Headstash potency and flavor. A mouth watering range of smells will be expressed from the plants: skunk, gasoline, pepper, vanilla cake frosting, and fermenting liqueur. Phenotypically, this hybrid doesn’t have all the hues you would see in other cookie hybrids but has all the other qualities you could ask for. ______ Dosidos x Key Lime Pie Type: Hybrid Finish time indoor / outdoor: 9 - 10 / October 5th - October 18th Yield: Medium Terp profile: Exotic Dosidos is another award winning strain that has quickly become a popular variety, that crossed with Key Lime Pie gives the strain some much desired improvements. Yield has been increased, structure improved, and all the no- ceiling potency of the Dosidos is still there. Terpene profile on these plants has been broadened and they exhibit a variation of aromas including burnt rubber, vanilla, pinesol, gasoline, and hints of papaya. Plants also can turn a dark purple hue on some phenotypes. ______ Gelato #45 x Key Lime Pie Type: Indica Finish time indoor / outdoor: 8 - 9 / September 28th - October 5th Yield: High Terp profile: Exotic An indica dominant cross which produces bright orange pistils with white resin that coats deep purple hued flowers. Expect phenotypes that will produce large flowers with short node spacing and a medium stature. -
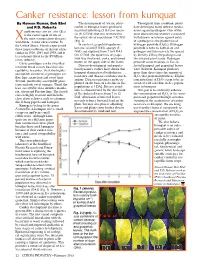
Canker Resistance: Lesson from Kumquat by Naveen Kumar, Bob Ebel the Development of Asiatic Citrus Throughout Their Evolution, Plants and P.D
Canker resistance: lesson from kumquat By Naveen Kumar, Bob Ebel The development of Asiatic citrus Throughout their evolution, plants and P.D. Roberts canker in kumquat leaves produced have developed many defense mecha- anthomonas citri pv. citri (Xcc) localized yellowing (5 DAI) or necro- nisms against pathogens. One of the is the causal agent of one of sis (9-12 DAI) that was restricted to most characteristic features associated the most serious citrus diseases the actual site of inoculation 7-12 DAI with disease resistance against entry X (Fig. 2). of a pathogen is the production of worldwide, Asiatic citrus canker. In the United States, Florida experienced In contrast, grapefruit epidermis hydrogen peroxide (H2O2). Hydrogen three major outbreaks of Asiatic citrus became raised (5 DAI), spongy (5 peroxide is toxic to both plant and canker in 1910, 1984 and 1995, and it DAI) and ruptured from 7 to 8 DAI. pathogen and thus restricts the spread is a constant threat to the $9 billion On 12 DAI, the epidermis of grape- by directly killing the pathogen and citrus industry. fruit was thickened, corky, and turned the infected plant tissue. Hydrogen Citrus genotypes can be classified brown on the upper side of the leaves. peroxide concentrations in Xcc-in- into four broad classes based on sus- Disease development and popula- fected kumquat and grapefruit leaves ceptibility to canker. First, the highly- tion dynamics studies have shown that were different. Kumquat produces susceptible commercial genotypes are kumquat demonstrated both disease more than three times the amount of Key lime, grapefruit and sweet lime. -

Citrus from Seed?
Which citrus fruits will come true to type Orogrande, Tomatera, Fina, Nour, Hernandina, Clementard.) from seed? Ellendale Tom McClendon writes in Hardy Citrus Encore for the South East: Fortune Fremont (50% monoembryonic) “Most common citrus such as oranges, Temple grapefruit, lemons and most mandarins Ugli Umatilla are polyembryonic and will come true to Wilking type. Because most citrus have this trait, Highly polyembryonic citrus types : will mostly hybridization can be very difficult to produce nucellar polyembryonic seeds that will grow true to type. achieve…. This unique characteristic Citrus × aurantiifolia Mexican lime (Key lime, West allows amateurs to grow citrus from seed, Indian lime) something you can’t do with, say, Citrus × insitorum (×Citroncirus webberii) Citranges, such as Rusk, Troyer etc. apples.” [12*] Citrus × jambhiri ‘Rough lemon’, ‘Rangpur’ lime, ‘Otaheite’ lime Monoembryonic (don’t come true) Citrus × limettioides Palestine lime (Indian sweet lime) Citrus × microcarpa ‘Calamondin’ Meyer Lemon Citrus × paradisi Grapefruit (Marsh, Star Ruby, Nagami Kumquat Redblush, Chironja, Smooth Flat Seville) Marumi Kumquat Citrus × sinensis Sweet oranges (Blonde, navel and Pummelos blood oranges) Temple Tangor Citrus amblycarpa 'Nasnaran' mandarin Clementine Mandarin Citrus depressa ‘Shekwasha’ mandarin Citrus karna ‘Karna’, ‘Khatta’ Poncirus Trifoliata Citrus kinokuni ‘Kishu mandarin’ Citrus lycopersicaeformis ‘Kokni’ or ‘Monkey mandarin’ Polyembryonic (come true) Citrus macrophylla ‘Alemow’ Most Oranges Citrus reshni ‘Cleopatra’ mandarin Changshou Kumquat Citrus sunki (Citrus reticulata var. austera) Sour mandarin Meiwa Kumquat (mostly polyembryonic) Citrus trifoliata (Poncirus trifoliata) Trifoliate orange Most Satsumas and Tangerines The following mandarin varieties are polyembryonic: Most Lemons Dancy Most Limes Emperor Grapefruits Empress Tangelos Fairchild Kinnow Highly monoembryonic citrus types: Mediterranean (Avana, Tardivo di Ciaculli) Will produce zygotic monoembryonic seeds that will not Naartje come true to type. -

Generation of Sexual and Somatic Hybrids in Acid Citrus Fruits
GENERATION OF SEXUAL AND SOMATIC HYBRIDS IN ACID CITRUS FRUITS By ZENAIDA JOSEFINA VILORIA VILLALOBOS A DISSERTATION PRESENTED TO THE GRADUATE SCHOOL OF THE UNIVERSITY OF FLORIDA IN PARTIAL FULFILLMENT OF THE REQUIREMENTS FOR THE DEGREE OF DOCTOR OF PHILOSOPHY UNIVERSITY OF FLORIDA 2003 Copyright 2003 by Zenaida Josefina Viloria Villalobos This dissertation is dedicated to my darling mother Olivia and to the memory of my beloved father Dimas, and to my sisters Celina, Doris, Celmira, and Olivia, and brothers Dimas, Silfredo and Alejandro, with love. ACKNOWLEDGMENTS This work was completed with the generous collaboration of many people to whom I will always be grateful. First I wish to thank my supervisor Dr. Jude Grosser, for his guidance, suggestions, and financial assistance during the last period of my studies. I also want to thank the University of Zulia and Fondo Nacional de Ciencias, Tecnologia e Innovation for giving me the opportunity to do my doctoral studies. I thank very much Dr. Renee Goodrich, Dr. Frederick Gmitter, Dr. Michael Kane and Dr. Dennis Gray for being members of my committee and for their contributions to this work. Thanks go to Dr. Glem Wright (University of Arizona) for making it possible to generate more lemon progenies in this study. I appreciate very much the supervision and help in completing the canker screening study from Dr. Graham, Diana Drouillard and Diane Bright. I thank very much Dr. Ramon Littell and Belkys Bracho for their assistance on the statistical analysis of my experiments. Thanks go to the Division of Plant Industry (Lake Alfred, FL), particularly to Mrs. -

Craft Cocktails
Craft Cocktails Horseshoes & Hand-Grenades Aviation Vodka, Cointreau, Strawberry Syrup, Lime Juice, Gin, Crème de Violette, Luxardo Cherry, Simple Grapefruit Juice, Basil Syrup, Lemon Juice $10 $10 Slippery Slope Sazerac Vodka, Pimm’s#1, Lemon Juice, Strawberry Syrup, Bulleit Rye, Absinthe, Sugar, Peychaud’s Bitters Pineapple Juice, Plum & Peychaud’s Bitters, $10 Egg White $9 Old Fashioned Spontaneously Horny Bulleit Bourbon, Sugar, Bitters, Orange Twist $12 Vodka, St. Germain, Blackberry Syrup, Lemon Juice, Mint Gotham Sour $10 Bulleit Bourbon, Lemon Juice, Simple Syrup, Egg Rusty Pear White, Red Wine Float $10 Absolut Pear, Domaine de Canton, Lemon Juice, Simple Syrup Maxwell’s Silver Hammer $9 Blanco Tequila, Cucumber, Pineapple Juice, Lime Adelaide Juice, Agave Nectar, Cointreau, Salted Rim $11 Gin, St. Germain, Lime Juice, Soda Water, Cucumber, Mint Lavender Love Potion #9 $10 Crème de Violette, St. Germain, Lime Juice, Joan of Arc Prosecco, Mint $12 Gin, Egg White, Lemon Juice, Simple Syrup, Raspberry Syrup, Prosecco, Mint Caipirinha $10 Cachaça Rum, Luxardo Maraschino, Simple Syrup, Lime Juice Smoking Gun $8 Bulleit Bourbon, Maple Syrup, Bitters, Applewood Smoke Dark & Stormy $12 Meyers Rum, Domaine de Canton, Ginger Beer, Fig Manhattan Lime $8 House Fig Whiskey, Sweet Vermouth, Luxardo Cherry, Candied Bacon Key Lime Martini $14 Vanilla Vodka, Pineapple Juice, Lime Juice, El Chapo Coconut Cream, Cookie Crumble Rim $10 Blanco Tequila, Cointreau, Lime Juice, Pineapple Juice, Strawberry Syrup, Jalapenos, The Catalyst Cilantro, Salted Rim Parfait Amour, St. Germain, Lemon Juice, $12 Lemon-Lime Soda, Prosecco, Luxardo Cherry TR Smash $9 Templeton Rye, Lemon, Mint, Honey French 75 $10 Gin, Lemon Juice, Simple Syrup, Prosecco $9 Don’t Forget to Check the Board for the Feature Cocktail and Martini . -

New and Noteworthy Citrus Varieties Presentation
New and Noteworthy Citrus Varieties Citrus species & Citrus Relatives Hundreds of varieties available. CITRON Citrus medica • The citron is believed to be one of the original kinds of citrus. • Trees are small and shrubby with an open growth habit. The new growth and flowers are flushed with purple and the trees are sensitive to frost. • Ethrog or Etrog citron is a variety of citron commonly used in the Jewish Feast of Tabernacles. The flesh is pale yellow and acidic, but not very juicy. The fruits hold well on the tree. The aromatic fruit is considerably larger than a lemon. • The yellow rind is glossy, thick and bumpy. Citron rind is traditionally candied for use in holiday fruitcake. Ethrog or Etrog citron CITRON Citrus medica • Buddha’s Hand or Fingered citron is a unique citrus grown mainly as a curiosity. The six to twelve inch fruits are apically split into a varying number of segments that are reminiscent of a human hand. • The rind is yellow and highly fragrant at maturity. The interior of the fruit is solid rind with no flesh or seeds. • Fingered citron fruits usually mature in late fall to early winter and hold moderately well on the tree, but not as well as other citron varieties. Buddha’s Hand or Fingered citron NAVEL ORANGES Citrus sinensis • ‘Washington navel orange’ is also known • ‘Lane Late Navel’ was the first of a as the Bahia. It was imported into the number of late maturing Australian United States in 1870. navel orange bud sport selections of Washington navel imported into • These exceptionally delicious, seedless, California. -

Citrus Trees Grow Very Well in the Sacramento Valley!
Citrus! Citrus trees grow very well in the Sacramento Valley! They are evergreen trees or large shrubs, with wonderfully fragrant flowers and showy fruit in winter. There are varieties that ripen in nearly every season. Citrus prefer deep, infrequent waterings, regular fertilizer applications, and may need protection from freezing weather. We usually sell citrus on rootstocks that make them grow more slowly, so we like to call them "semi-dwarf". We can also special-order most varieties on rootstocks that allow them to grow larger. Citrus size can be controlled by pruning. The following citrus varieties are available from the Redwood Barn Nursery, and are recommended for our area unless otherwise noted in the description. Oranges Robertson Navel Best selling winter-ripening variety. Early and heavy bearing. Cultivar of Washington Navel. Washington Navel California's famous winter-ripening variety. Fruit ripens in ten months. Jaffa (Shamouti) Fabled orange from Middle East. Very few seeds. spring to summer ripening. Good flavor. Trovita Spring ripening. Good in many locations from coastal areas to desert. Few seeds, heavy producer, excellent flavor. Valencia Summer-ripening orange for juicing or eating. Fifteen months to ripen. Grow your own orange juice. Seville Essential for authentic English marmalade. Used fresh or dried in Middle Eastern cooking. Moro Deep blood coloration, almost purple-red, even in California coastal areas. Very productive, early maturity, distinctive aroma, exotic berry-like flavor. Sanquinella A deep blood red juice and rind. Tart, spicy flavor. Stores well on tree. Mandarins / Tangerines Dancy The best-known Mandarin type. On fruit stands at Christmas time. -

Mexican Lime
MEXICAN LIME Commercial citrus production in the Yuma area is devoted primarily to tangelo/tangerines, a distant second to desert lemons, the major citrus fruit grown in the area. Mexican limes, however, fit into a local niche market, with the small acreage grown essentially for local markets. · Of the two acid, or sour, limes in world trade, the one longest known and most widely cultivated is the Mexican lime (Citrus aurantifoli), and many often refer the tangy fruit merely as "lime". · The Mexican lime tree is exceptionally vigorous; may be shrubby and range from 6 1/2 to 13 feet high, with many slender, spreading branches, and usually with numerous, very sharp spines to 3/8 inch long. The lime fruits are borne singly or in 2's or 3's (or sometimes large clusters), at the twig tips. The pulp of the Mexican lime is greenish-yellow and the fruits are quite juicy, very acid and flavorful, with few or many small seeds which are also green in color. · The Mexican lime, because of its special bouquet and unique flavor, is ideal for serving in half as a garnish and flavoring for fish and meats, for adding zest to cold drinks, and for making limeade. Commercially bottled lime juice is prized the world over for use in mixed alcoholic drinks. · Limes are a very juicy citrus fruit. In fact, it is calculated that 2,200 lbs of fruit yields 1,058 pounds of juice. · Mexican limes are often made into jam, jelly and marmalade. They are also pickled by first making 4 incisions at the top of the fruit and covering the fruits with salt, and later preserv- ing them in vinegar. -

Citrus Offers Year-Round Options
for the Gardener Citrus Offers Year-Round Options he genus Citrus is undoubtedly the premier genus in the Rutaceae (Rue) TABLE 1. GENUS CITRUS: PROMINENT SPECIES AND PLACES OF ORIGIN, FROM LEAST COLD HARDY TO MOST COLD HARDY family. This family features 150 genera T 32ºF* Citrus medica India Citron/Etrog, Buddha’s Hand Fruit and 1600 species and consists largely 32ºF C. aurantiifolia India, S.E. Asia Limes: Mexican, Bearss, of evergreen shrubs and trees from the Key Lime, Tahitian Lime Mediterranean, subtropical, semitropical 28ºF C. limon S.E. Asia Lemons or limonia and tropical zones of the world. Table 1 26ºF C. paradisi Carribean Grapefruit: natural hybrid between (at right) lists the prominent species of lemon and pummelo the genus Citrus (and Fortunella), their 24ºF C. maxima Malaysian peninsula Pummelo hardiness, and place of origin. Among or grandis the other significant food and ornamental 24ºF C. paradisi (grapefruit) x Tangelo: bred cross of grapefruit genera in the family are – reticulata (Mandarin) and Mandarin Choisya (Mock orange) 24ºF C. sinensis S. E. China, Vietnam Oranges: sweet, sour, blood 24ºF C. nobilis (King) Florida Oranges: Temple, sweet Correa (Coral bells) reticulata x sinensis Dictamnus (Burning bush) 22ºF C. reticulata S.E. Asia Mandarins, Tangerines Casimiroa (White zapote) 22ºF C. limon cv. ‘Improved Meyer Lemon’ Bred hybrid cross, orange x lemon Fortunella (Kumquats and their 20ºF C. mitis Phillipines Calamondin 18ºF Fortunella S.E. China Kumquat, Limequat, Orangequat, hybrid forms) -20ºF margarita and hybrids Citrangequat Poncirus (Trifoliate orange and * approximate temperature at which frost damage begins to occur (flowers are flying dragon) the most cold-sensitive, followed by fruit, leaves, and wood) Ruta (Rue) Members of the Rue family feature strong oil glands in both the leaves and fruit.