New England Seaweed Culture Handbook Sarah Redmond University of Connecticut - Stamford, [email protected]
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-
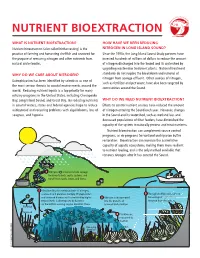
Nutrient Bioextraction L
soun nd d s sla tu i d g y n o NUTRIENT BIOEXTRACTION l WHAT IS NUTRIENT BIOEXTRACTION? HOW HAVE WE BEEN REDUCING Nutrient bioextraction (also called bioharvesting) is the NITROGEN IN LONG ISLAND SOUND? practice of farming and harvesting shellfish and seaweed for Since the 1990s, the Long Island Sound Study partners have the purpose of removing nitrogen and other nutrients from invested hundreds of millions of dollars to reduce the amount natural water bodies. of nitrogen discharged into the Sound and its watershed by upgrading wastewater treatment plants. National treatment WHY DO WE CARE ABOUT NITROGEN? standards do not require the breakdown and removal of nitrogen from sewage effluent. Other sources of nitrogen, Eutrophication has been identified by scientists as one of such as fertilizer and pet waste, have also been targeted by the most serious threats to coastal environments around the communities around the Sound. world. Reducing nutrient inputs is a top priority for many estuary programs in the United States, including Chesapeake Bay, Long Island Sound, and Great Bay. By reducing nutrients WHY DO WE NEED NUTRIENT BIOEXTRACTION? in coastal waters, states and federal agencies hope to reduce Efforts to control nutrient sources have reduced the amount widespread and recurring problems with algal blooms, loss of of nitrogen entering the Sound each year. However, changes seagrass, and hypoxia. in the Sound and its watershed, such as wetland loss and decreased populations of filter feeders, have diminished the capacity of the system to naturally process and treat nutrients. Nutrient bioextraction can complement source control programs, as do programs for wetland and riparian buffer restoration. -

RED ALGAE · RHODOPHYTA Rhodophyta Are Cosmopolitan, Found from the Artic to the Tropics
RED ALGAE · RHODOPHYTA Rhodophyta are cosmopolitan, found from the artic to the tropics. Although they grow in both marine and fresh water, 98% of the 6,500 species of red algae are marine. Most of these species occur in the tropics and sub-tropics, though the greatest number of species is temperate. Along the California coast, the species of red algae far outnumber the species of green and brown algae. In temperate regions such as California, red algae are common in the intertidal zone. In the tropics, however, they are mostly subtidal, growing as epiphytes on seagrasses, within the crevices of rock and coral reefs, or occasionally on dead coral or sand. In some tropical waters, red algae can be found as deep as 200 meters. Because of their unique accessory pigments (phycobiliproteins), the red algae are able to harvest the blue light that reaches deeper waters. Red algae are important economically in many parts of the world. For example, in Japan, the cultivation of Pyropia is a multibillion-dollar industry, used for nori and other algal products. Rhodophyta also provide valuable “gums” or colloidal agents for industrial and food applications. Two extremely important phycocolloids are agar (and the derivative agarose) and carrageenan. The Rhodophyta are the only algae which have “pit plugs” between cells in multicellular thalli. Though their true function is debated, pit plugs are thought to provide stability to the thallus. Also, the red algae are unique in that they have no flagellated stages, which enhance reproduction in other algae. Instead, red algae has a complex life cycle, with three distinct stages. -

Red Algae (Bangia Atropurpurea) Ecological Risk Screening Summary
Red Algae (Bangia atropurpurea) Ecological Risk Screening Summary U.S. Fish & Wildlife Service, February 2014 Revised, March 2016, September 2017, October 2017 Web Version, 6/25/2018 1 Native Range and Status in the United States Native Range From NOAA and USGS (2016): “Bangia atropurpurea has a widespread amphi-Atlantic range, which includes the Atlantic coast of North America […]” Status in the United States From Mills et al. (1991): “This filamentous red alga native to the Atlantic Coast was observed in Lake Erie in 1964 (Lin and Blum 1977). After this sighting, records for Lake Ontario (Damann 1979), Lake Michigan (Weik 1977), Lake Simcoe (Jackson 1985) and Lake Huron (Sheath 1987) were reported. It has become a major species of the littoral flora of these lakes, generally occupying the littoral zone with Cladophora and Ulothrix (Blum 1982). Earliest records of this algae in the basin, however, go back to the 1940s when Smith and Moyle (1944) found the alga in Lake Superior tributaries. Matthews (1932) found the alga in Quaker Run in the Allegheny drainage basin. Smith and 1 Moyle’s records must have not resulted in spreading populations since the alga was not known in Lake Superior as of 1987. Kishler and Taft (1970) were the most recent workers to refer to the records of Smith and Moyle (1944) and Matthews (1932).” From NOAA and USGS (2016): “Established where recorded except in Lake Superior. The distribution in Lake Simcoe is limited (Jackson 1985).” From Kipp et al. (2017): “Bangia atropurpurea was first recorded from Lake Erie in 1964. During the 1960s–1980s, it was recorded from Lake Huron, Lake Michigan, Lake Ontario, and Lake Simcoe (part of the Lake Ontario drainage). -

The Global Dispersal of the Non-Endemic Invasive Red Alga Gracilariavermiculophylla in the Ecosystems of the Euro-Asia Coastal W
Review Article Oceanogr Fish Open Access J Volume 8 Issue 1 - July 2018 Copyright © All rights are reserved by Vincent van Ginneken DOI: 10.19080/OFOAJ.2018.08.555727 The Global Dispersal of the Non-Endemic Invasive Red Alga Gracilaria vermiculophylla in the Ecosystems of the Euro-Asia Coastal Waters Including the Wadden Sea Unesco World Heritage Coastal Area: Awful or Awesome? Vincent van Ginneken* and Evert de Vries Bluegreentechnologies, Heelsum, Netherlands Submission: September 05, 2017; Published: July 06, 2018 Corresponding author: Vincent van Ginneken, Bluegreentechnologies, Heelsum, Netherlands, Email: Abstract Gracilaria vermiculophylla (Ohmi) Papenfu ß 1967 (Rhodophyta, Gracilariaceae) is a red alga and was originally described in Japan in 1956 as Gracilariopsis vermiculophylla G. vermiculophylla is primarily used as a precursor for agar, which is widely used in the pharmaceutical and food industries. It has been introduced to the East . It is thought to be native and widespread throughout the Northwest Pacific Ocean. temperature) and can grow in an extremely wide variety of conditions; factors which contribute to its invasiveness. It invades estuarine areas Pacific, the West Atlantic and the East Atlantic, where it rapidly colonizes new environments. It is highly tolerant of stresses (nutrient, salinity, invaded: Atlantic, North Sea, Mediterranean and Baltic Sea. The Euro-Asian brackish Black-Sea have not yet been invaded but are very vulnerable towhere intense it out-competes invasion with native G. vermiculophylla algae species and modifies environments. The following European coastal and brackish water seas are already G. vermiculophylla among the most potent invaders out of 114 non-indigenous because they macro-algae are isolated species from indirect Europe. -

(Rhodophyta, Gracilariales) in Hog Island Bay, Virginia: a Cryptic Alien and Invasive Macroalga and Taxonomic Correction1
J. Phycol. 42, 139–141 (2005) r 2005 Phycological Society of America DOI: 10.1111/j.1529-8817.2005.00160.x NOTE GRACILARIA VERMICULOPHYLLA (RHODOPHYTA, GRACILARIALES) IN HOG ISLAND BAY, VIRGINIA: A CRYPTIC ALIEN AND INVASIVE MACROALGA AND TAXONOMIC CORRECTION1 Mads Solgaard Thomsen2 Department of Applied Sciences, Auckland University of Technology, Auckland, New Zealand Carlos Frederico Deluqui Gurgel, Suzanne Fredericq Department of Biology, University of Louisiana at Lafayette, P.O. Box 42451, Lafayette, Louisiana 70504-2451, USA and Karen J. McGlathery Department of Environmental Sciences, Clark Hall, University of Virginia, Charlottesville, Virginia 22903, USA Gracilaria in Virginia, USA, is abundant and Lachlan and Bird 1986). Unfortunately, in many cases, composed of thalli either having relatively flat or Gracilaria species are difficult to identify based on mor- cylindrical branches. These two morphologies were phological features (Oliveira et al. 2000, Gurgel and referred to previously as G. foliifera (Forsska˚l) Fredericq 2004). Given these difficulties and new pos- Bgesen and G. verrucosa (Hudson) Papenfuss. sibilities of accurate identification by molecular biology However, G. verrucosa is regarded an invalid techniques, Gracilaria sensu lato species are regularly name, and the flat specimens are now referred to as changing taxonomic status (Bird and Rice 1990, Bell- G. tikvahiae McLachlan. This has created confusion orin et al. 2002, Gurgel and Fredericq 2004, Gurgel about the nomenclature of Gracilaria from this re- et al. 2004). gion. Here we document that the cylindrical form Gracilaria is a particularly important genus in Vir- that dominates Hog Island Bay, Virginia, is ginia, USA, where it is abundant in lagoons and estu- G. -

Modelling Potential Production and Environmental Effects Of
Biogeosciences Discuss., https://doi.org/10.5194/bg-2017-195 Manuscript under review for journal Biogeosciences Discussion started: 14 June 2017 c Author(s) 2017. CC BY 3.0 License. Modelling potential production and environmental effects of macroalgae farms in UK and Dutch coastal waters Johan van der Molen1,2, Piet Ruardij2, Karen Mooney4, Philip Kerrison5, Nessa E. O'Connor4, Emma Gorman4, Klaas Timmermans3, Serena Wright1, Maeve Kelly5, Adam D. Hughes5, Elisa Capuzzo1 5 1The Centre for Environment, Fisheries and Aquaculture Science (Cefas), Lowestoft, NR33 0HT, UK 2NIOZ Royal Netherlands Institute for Sea Research, Dept. of Coastal Systems and Utrecht University, Den Burg, 1797 SZ, The Netherlands 3NIOZ Royal Netherlands Institute for Sea Research, Dept. of Estuarine and Delta Systems and Utrecht University, Yerseke, 4401 NT, The Netherlands 10 4Queen’s University, Belfast, BT7 1NN, UK 5The Scottish Association for Marine Science (SAMS), Oban, PA37 1QA, UK Correspondence to: Johan van der Molen ([email protected], [email protected]) Abstract. There is increasing interest in macroalgae farming in European waters for a range of applications, including food, chemical extraction and as biofuels. This study uses a 3D numerical model of hydrodynamics and biogeochemistry to 15 investigate potential production and environmental effects of macroalgae farming in UK and Dutch coastal waters. The model included four experimental farms in different coastal settings in Strangford Lough (Northern Ireland), in Sound of Kerrera and Lynn of Lorne (northwest Scotland), and in the Rhine Plume (The Netherlands), as well as a hypothetical large- scale farm off the UK north Norfolk coast. -

Marlin Marine Information Network Information on the Species and Habitats Around the Coasts and Sea of the British Isles
MarLIN Marine Information Network Information on the species and habitats around the coasts and sea of the British Isles Sugar kelp (Saccharina latissima) MarLIN – Marine Life Information Network Biology and Sensitivity Key Information Review Nicola White & Charlotte Marshall 2007-09-06 A report from: The Marine Life Information Network, Marine Biological Association of the United Kingdom. Please note. This MarESA report is a dated version of the online review. Please refer to the website for the most up-to-date version [https://www.marlin.ac.uk/species/detail/1375]. All terms and the MarESA methodology are outlined on the website (https://www.marlin.ac.uk) This review can be cited as: White, N. & Marshall, C.E. 2007. Saccharina latissima Sugar kelp. In Tyler-Walters H. and Hiscock K. (eds) Marine Life Information Network: Biology and Sensitivity Key Information Reviews, [on-line]. Plymouth: Marine Biological Association of the United Kingdom. DOI https://dx.doi.org/10.17031/marlinsp.1375.1 The information (TEXT ONLY) provided by the Marine Life Information Network (MarLIN) is licensed under a Creative Commons Attribution-Non-Commercial-Share Alike 2.0 UK: England & Wales License. Note that images and other media featured on this page are each governed by their own terms and conditions and they may or may not be available for reuse. Permissions beyond the scope of this license are available here. Based on a work at www.marlin.ac.uk (page left blank) Date: 2007-09-06 Sugar kelp (Saccharina latissima) - Marine Life Information Network See online review for distribution map Buoy line with Saccharina latissima. -

27April12acquatic Plants
International Plant Protection Convention Protecting the world’s plant resources from pests 01 2012 ENG Aquatic plants their uses and risks Implementation Review and Support System Support and Review Implementation A review of the global status of aquatic plants Aquatic plants their uses and risks A review of the global status of aquatic plants Ryan M. Wersal, Ph.D. & John D. Madsen, Ph.D. i The designations employed and the presentation of material in this information product do not imply the expression of any opinion whatsoever on the part of the Food and Agriculture Organization of the United Nations (FAO) concerning the legal or development status of any country, territory, city or area or of its authorities, or concerning the delimitation of its frontiers or boundaries. The mention of speciic companies or products of manufacturers, whether or not these have been patented, does not imply that these have been endorsed or recommended by FAO in preference to others of a similar nature that are not mentioned.All rights reserved. FAO encourages reproduction and dissemination of material in this information product. Non-commercial uses will be authorized free of charge, upon request. Reproduction for resale or other commercial purposes, including educational purposes, may incur fees. Applications for permission to reproduce or disseminate FAO copyright materials, and all queries concerning rights and licences, should be addressed by e-mail to [email protected] or to the Chief, Publishing Policy and Support Branch, Ofice of Knowledge Exchange, -

Kelp Cultivation Effectively Improves Water Quality and Regulates Phytoplankton Community in a Turbid, Highly Eutrophic Bay
Science of the Total Environment 707 (2020) 135561 Contents lists available at ScienceDirect Science of the Total Environment journal homepage: www.elsevier.com/locate/scitotenv Kelp cultivation effectively improves water quality and regulates phytoplankton community in a turbid, highly eutrophic bay Zhibing Jiang a,b,d, Jingjing Liu a,ShangluLic,YueChena,PingDua,b,YuanliZhua,YiboLiaoa,d, Quanzhen Chen a, Lu Shou a, Xiaojun Yan e, Jiangning Zeng a,⁎, Jianfang Chen a,d a Key Laboratory of Marine Ecosystem and Biogeochemistry, State Oceanic Administration & Second Institute of Oceanography, Ministry of Natural Resources, Hangzhou, China b Function Laboratory for Marine Fisheries Science and Food Production Processes, Pilot National Laboratory for Marine Science and Technology (Qingdao), Qingdao, China c Marine Monitoring and Forecasting Center of Zhejiang Province, Hangzhou, China d State Key Laboratory of Satellite Ocean Environment Dynamics, Second Institute of Oceanography, Ministry of Natural Resources, Hangzhou, China e Key Laboratory of Applied Marine Biotechnology, Ministry of Education, Marine College of Ningbo University, Ningbo, China HIGHLIGHTS GRAPHICAL ABSTRACT • Kelp farming alleviated eutrophication and acidification. • Kelp farming greatly relieved light limi- tation and increased phytoplankton bio- mass. • Kelp farming appreciably enhanced phytoplankton diversity. • Kelp farming reduced the dominance of dinoflagellate Prorocentrum minimum. • Phytoplankton community differed sig- nificantly between the kelp farm and control area. article info abstract Article history: Coastal eutrophication and its associated harmful algal blooms have emerged as one of the most severe environ- Received 19 September 2019 mental problems worldwide. Seaweed cultivation has been widely encouraged to control eutrophication and Received in revised form 11 November 2019 algal blooms. Among them, cultivated kelp (Saccharina japonica) dominates primarily by production and area. -

The Life Cycle and Genetic Structure of the Red Alga Furcellaria Lumbricalis on a Salinity Gradient
WALTER AND ANDRÉE DE NOTTBECK FOUNDATION SCIENTIFIC REPORTS View metadata, citation and similar papers at core.ac.uk brought to you by CORE No. 33 provided by Helsingin yliopiston digitaalinen arkisto The life cycle and genetic structure of the red alga Furcellaria lumbricalis on a salinity gradient KIRSI KOSTAMO Academic dissertation in Ecology, to be presented, with the permission of the Faculty of Biosciences of the University of Helsinki, for public criticism in the Auditorium 1041, Biocenter 2, University of Helsinki, Viikinkaari 5, Helsinki, on January 25th, 2008, at 12 noon. HELSINKI 2008 This thesis is based on the following papers, which will be referred to in the text by their Roman numerals: I. Kostamo, K. & Mäkinen, A. 2006: Observations on the mode and seasonality of reproduction in Furcellaria lumbricalis (Gigartinales, Rhodophyta) populations in the northern Baltic Sea. – Bot. Mar. 49: 304-309. II Korpelainen, H., Kostamo, K. & Virtanen, V. 2007: Microsatellite marker identifi cation using genome screening and restriction-ligation. – BioTechniques 42: 479-486. III Kostamo, K., Korpelainen, H., Maggs, C. A. & Provan, J. 2007: Genetic variation among populations of the red alga Furcellaria lumbricalis in northern Europe. – Manuscript. IV Kostamo, K. & Korpelainen, H. 2007: Clonality and small-scale genetic diversity within populations of the red alga Furcellaria lumbricalis (Rhodophyta) in Ireland and in the northern Baltic Sea. – Manuscript. Paper I is printed with permission from Walter de Gruyter Publishers and paper II with permission from BioTechniques. Reviewers: Dr. Elina Leskinen Department of Biological and Environmental Sciences University of Helsinki Finland Prof. Kerstin Johannesson Tjärnö Marine Biological Laboratory University of Gothenburg Sweden Opponent: Dr. -

Download PDF Version
MarLIN Marine Information Network Information on the species and habitats around the coasts and sea of the British Isles Dabberlocks (Alaria esculenta) MarLIN – Marine Life Information Network Biology and Sensitivity Key Information Review Dr Harvey Tyler-Walters 2008-05-29 A report from: The Marine Life Information Network, Marine Biological Association of the United Kingdom. Please note. This MarESA report is a dated version of the online review. Please refer to the website for the most up-to-date version [https://www.marlin.ac.uk/species/detail/1291]. All terms and the MarESA methodology are outlined on the website (https://www.marlin.ac.uk) This review can be cited as: Tyler-Walters, H., 2008. Alaria esculenta Dabberlocks. In Tyler-Walters H. and Hiscock K. (eds) Marine Life Information Network: Biology and Sensitivity Key Information Reviews, [on-line]. Plymouth: Marine Biological Association of the United Kingdom. DOI https://dx.doi.org/10.17031/marlinsp.1291.1 The information (TEXT ONLY) provided by the Marine Life Information Network (MarLIN) is licensed under a Creative Commons Attribution-Non-Commercial-Share Alike 2.0 UK: England & Wales License. Note that images and other media featured on this page are each governed by their own terms and conditions and they may or may not be available for reuse. Permissions beyond the scope of this license are available here. Based on a work at www.marlin.ac.uk (page left blank) Date: 2008-05-29 Dabberlocks (Alaria esculenta) - Marine Life Information Network See online review for distribution map Exposed sublittoral fringe bedrock with Alaria esculenta, Isles of Scilly. -

Chemical Composition and Potential Practical Application of 15 Red Algal Species from the White Sea Coast (The Arctic Ocean)
molecules Article Chemical Composition and Potential Practical Application of 15 Red Algal Species from the White Sea Coast (the Arctic Ocean) Nikolay Yanshin 1, Aleksandra Kushnareva 2, Valeriia Lemesheva 1, Claudia Birkemeyer 3 and Elena Tarakhovskaya 1,4,* 1 Department of Plant Physiology and Biochemistry, Faculty of Biology, St. Petersburg State University, 199034 St. Petersburg, Russia; [email protected] (N.Y.); [email protected] (V.L.) 2 N. I. Vavilov Research Institute of Plant Industry, 190000 St. Petersburg, Russia; [email protected] 3 Faculty of Chemistry and Mineralogy, University of Leipzig, 04103 Leipzig, Germany; [email protected] 4 Vavilov Institute of General Genetics RAS, St. Petersburg Branch, 199034 St. Petersburg, Russia * Correspondence: [email protected] Abstract: Though numerous valuable compounds from red algae already experience high demand in medicine, nutrition, and different branches of industry, these organisms are still recognized as an underexploited resource. This study provides a comprehensive characterization of the chemical composition of 15 Arctic red algal species from the perspective of their practical relevance in medicine and the food industry. We show that several virtually unstudied species may be regarded as promis- ing sources of different valuable metabolites and minerals. Thus, several filamentous ceramialean algae (Ceramium virgatum, Polysiphonia stricta, Savoiea arctica) had total protein content of 20–32% of dry weight, which is comparable to or higher than that of already commercially exploited species Citation: Yanshin, N.; Kushnareva, (Palmaria palmata, Porphyra sp.). Moreover, ceramialean algae contained high amounts of pigments, A.; Lemesheva, V.; Birkemeyer, C.; macronutrients, and ascorbic acid. Euthora cristata (Gigartinales) accumulated free essential amino Tarakhovskaya, E.