Assessing New Methods for Psittacine Conservation at The
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

TAG Operational Structure
PARROT TAXON ADVISORY GROUP (TAG) Regional Collection Plan 5th Edition 2020-2025 Sustainability of Parrot Populations in AZA Facilities ...................................................................... 1 Mission/Objectives/Strategies......................................................................................................... 2 TAG Operational Structure .............................................................................................................. 3 Steering Committee .................................................................................................................... 3 TAG Advisors ............................................................................................................................... 4 SSP Coordinators ......................................................................................................................... 5 Hot Topics: TAG Recommendations ................................................................................................ 8 Parrots as Ambassador Animals .................................................................................................. 9 Interactive Aviaries Housing Psittaciformes .............................................................................. 10 Private Aviculture ...................................................................................................................... 13 Communication ........................................................................................................................ -

Oceania Species ID Sheets
Species Identification Sheets for Protected Wildlife in Trade - Oceania - 3 Mark O’Shea 1 Mike McCoy © Phil Bender 5 Tony Whitaker © 2 4 Tony Whitaker © 6 WILDLIFE ENFORCEMENT GROUP (AGRICULTURE & FORESTRY · CONSERVATION · N. Z. CUSTOMS SERVICE) Numbered images above Crown Copyright: Department of Conservation Te Papa Atawhai. Photographers:1) Dick Veitch 1981, 2) Rod Morris 1984, 3) Gareth Rapley 2009, 4) Andrew Townsend 2000, 5) Paul Schilov 2001, 6) Dick Veitch 1979 Introduction Purpose of this resource: - Additional species that should be included in this booklet Wildlife trafficking is a large-scale multi-billion dollar industry worldwide. The illegal trade of - Sources of information, such as identification guides or reports, related to these wildlife has reached such prominence that it has the potential to devastate source populations species of wildlife, impacting on the integrity and productivity of ecosystems in providing food and - Domestic legislation regarding the regulation of trade in wildlife - Sources of photographs for identification purposes resources to the local economy. In order to protect these resources, legislation has been put in place to control the trade of wildlife in almost every country worldwide. Those assigned with - Details of wildlife seizures, including the smuggling methods enforcing these laws have the monumental task of identifying the exact species that are being traded, either as whole living plants or animals, as parts that are dried, fried or preserved, or as Any feedback can be provided directly to the Wildlife Enforcement Group: derivatives contained within commercial products. Stuart Williamson Senior Investigator, Wildlife Enforcement Group This booklet “Species Identification Sheets for Protected Species in Trade – Oceania” has been Customhouse, Level 6, 50 Anzac Avenue, Auckland, New Zealand developed to address the lack of resources, identified by customs agencies within Oceania, for Ph: +64 9 3596676, Fax: +64 9 3772534 identification of wildlife species in trade. -

The Following Is a Section of a Document Properly Cited As: Snyder, N., Mcgowan, P., Gilardi, J., and Grajal, A. (Eds.) (2000) P
The following is a section of a document properly cited as: Snyder, N., McGowan, P., Gilardi, J., and Grajal, A. (eds.) (2000) Parrots. Status Survey and Conservation Action Plan 2000–2004. IUCN, Gland, Switzerland and Cambridge, UK. x + 180 pp. © 2000 International Union for Conservation of Nature and Natural Resources and the World Parrot Trust It has been reformatted for ease of use on the internet . The resolution of the photographs is considerably reduced from the printed version. If you wish to purchase a printed version of the full document, please contact: IUCN Publications Unit 219c Huntingdon Road, Cambridge, CB3 0DL, UK. Tel: (44) 1223 277894 Fax: (44) 1223 277175 Email: [email protected] The World Parrot Trust World Parrot Trust UK World Parrot Trust USA Order on-line at: Glanmor House PO Box 353 www.worldparrottrust.org Hayle, Cornwall Stillwater, MN 55082 TR27 4HB, United Kingdom Tel: 651 275 1877 Tel: (44) 1736 753365 Fax: 651 275 1891 Fax (44) 1736 751028 Island Press Box 7, Covelo, California 95428, USA Tel: 800 828 1302, 707 983 6432 Fax: 707 983 6414 E-mail: [email protected] Order on line: www.islandpress.org The views expressed in this publication are those of the authors and do not necessarily reflect those of IUCN or the Species Survival Commission. Published by: IUCN, Gland, Switzerland, and Cambridge, UK. Copyright: © 2000 International Union for Conservation of Nature and Natural Resources and the World Parrot Trust Reproduction of this publication for educational and other non-commercial purposes is authorised without prior written permission from the copyright holders provided the source is fully acknowledged. -

Chapter 1: Introduction
The following is a section of a document properly cited as: Snyder, N., McGowan, P., Gilardi, J., and Grajal, A. (eds.) (2000) Parrots. Status Survey and Conservation Action Plan 2000–2004. IUCN, Gland, Switzerland and Cambridge, UK. x + 180 pp. © 2000 International Union for Conservation of Nature and Natural Resources and the World Parrot Trust It has been reformatted for ease of use on the internet . The resolution of the photographs is considerably reduced from the printed version. If you wish to purchase a printed version of the full document, please contact: IUCN Publications Unit 219c Huntingdon Road, Cambridge, CB3 0DL, UK. Tel: (44) 1223 277894 Fax: (44) 1223 277175 Email: [email protected] The World Parrot Trust World Parrot Trust UK World Parrot Trust USA Order on-line at: Glanmor House PO Box 353 www.worldparrottrust.org Hayle, Cornwall Stillwater, MN 55082 TR27 4HB, United Kingdom Tel: 651 275 1877 Tel: (44) 1736 753365 Fax: 651 275 1891 Fax (44) 1736 751028 Island Press Box 7, Covelo, California 95428, USA Tel: 800 828 1302, 707 983 6432 Fax: 707 983 6414 E-mail: [email protected] Order on line: www.islandpress.org The views expressed in this publication are those of the authors and do not necessarily reflect those of IUCN or the Species Survival Commission. Published by: IUCN, Gland, Switzerland, and Cambridge, UK. Copyright: © 2000 International Union for Conservation of Nature and Natural Resources and the World Parrot Trust Reproduction of this publication for educational and other non-commercial purposes is authorised without prior written permission from the copyright holders provided the source is fully acknowledged. -

Captive Wildlife Division 2 Restricted List
Restricted Listing – Division 2 Animals currently in the province that are listed in Division 2 are not considered overtly dangerous, but pose other risks. They may be held for the remaining life of the animal only if they were in a person’s possession prior to June 1, 2021, and if the ministry is notified by November 30, 2021. Item Class (Common Name) Scientific Name PART 1 AMPHIBIANS (CLASS AMPHIBIA) 1. Family Ambystomatidae Gray Tiger Salamander Ambystoma mavortium diaboli Barred Tiger Salamander Ambystoma mavortium mavortium Eastern Tiger Salamander Ambystoma tigrinum 2. Family Bufonidae Canadian Toad Anaxyrus hemiophrys Great Plains Toad Bufo cognatus 3. Family Dendrobatidae Rockstone Poison Dart Frog Dendrobates nubeculosus 4. Family Hylidae Gray Tree Frog Dryophytes versicolor Tiger Leg Frog Pithecopus hypochondrialis Boreal Chorus Frog Pseudacris maculata 5. Family Hynobiidae Tokyo Salamander Hynobius tokyoensis 6. Family Pipidae African Clawed Frog Xenopus laevis 7. Family Ranidae Northern Leopard Frog Lithobates pipiens Wood Frog Lithobates sylvaticus 8. Family Rhacophoridae Flying Foam Nest Frog Polypedates leucomystax Vietnam Flying Frog Rhacophorus calcaneus Item Class (Common Name) Scientific Name Indonesian Flying Frog Rhacophorus reinwardtii Mossy Frog Theloderma bicolor 9. Family Scaphiopodidae Plains Spadefoot Toad Spea bombifrons 10. Family Sirenidae Lesser Siren Siren intermedia Greater Siren Siren lacertina PART 2 BIRDS (CLASS AVES) 1. Family Corvidae American Crow Corvus brachyrhynchos Black-billed Magpie Pica hudsonia 2. Family Icteridae Red-winged Blackbird Agelaius phoeniceus Brewer’s Blackbird Euphagus cyanocephalus Brown-headed Cowbird Molothrus ater Common Grackle Quiscalus quiscula Yellow-headed Blackbird Xanthocephalus xanthocephalus 3. Family Passeridae House Sparrow Passer domesticus 4. Family Phasianidae Cheer Pheasant Catreus wallichii Brown-eared Pheasant Crossoptilon mantchuricum Himalayan Monal Lophophorus impejanus Edwards’s Pheasant Lophura edwardsi Swinhoe’s Pheasant Lophura swinhoii Elliot’s Pheasant Syrmaticus ellioti 5. -
The Extinction of the Carolina Parakeet and Multiple Dimensions of Global Parrot Biodiversity Kevin R
University of Connecticut OpenCommons@UConn Doctoral Dissertations University of Connecticut Graduate School 5-4-2017 The Extinction of the Carolina Parakeet and Multiple Dimensions of Global Parrot Biodiversity Kevin R. Burgio University of Connecticut - Storrs, [email protected] Follow this and additional works at: https://opencommons.uconn.edu/dissertations Recommended Citation Burgio, Kevin R., "The Extinction of the Carolina Parakeet and Multiple Dimensions of Global Parrot Biodiversity" (2017). Doctoral Dissertations. 1422. https://opencommons.uconn.edu/dissertations/1422 The Extinction of the Carolina Parakeet and Multiple Dimensions of Global Parrot Biodiversity Kevin Roy Burgio, PhD University of Connecticut, 2017 The study of the ecology of a species has traditionally ceased when that species goes extinct, despite the benefit to current and future generations of potential findings. We used the Carolina parakeet to develop a framework investigating the distributional limits, migratory habits, and extinction process as a means to recover important information. We developed a comprehensive database of every known occurrence of this iconic species. Using a combination of environmental niche modeling and extinction estimating analyses, our results demonstrate that the Carolina parakeet’s range was smaller than previously believed, the eastern and western subspecies occupied different niches with broad geographic separation, and that the western subspecies was a seasonal migrant while the eastern subspecies was not. We also found that it was likely habitat loss played a major role in their extinction. Our study highlights the importance of collecting occurrence data of extinct species and provides a framework for further investigations of other extinct species. Moreover, the recovery of lost autecological knowledge could benefit the conservation of other species currently in decline. -
Chapter 4 Australia, New Zealand, and the South-West Pacific
The following is a section of a document properly cited as: Snyder, N., McGowan, P., Gilardi, J., and Grajal, A. (eds.) (2000) Parrots. Status Survey and Conservation Action Plan 2000–2004. IUCN, Gland, Switzerland and Cambridge, UK. x + 180 pp. © 2000 International Union for Conservation of Nature and Natural Resources and the World Parrot Trust It has been reformatted for ease of use on the internet . The resolution of the photographs is considerably reduced from the printed version. If you wish to purchase a printed version of the full document, please contact: IUCN Publications Unit 219c Huntingdon Road, Cambridge, CB3 0DL, UK. Tel: (44) 1223 277894 Fax: (44) 1223 277175 Email: [email protected] The World Parrot Trust World Parrot Trust UK World Parrot Trust USA Order on-line at: Glanmor House PO Box 353 www.worldparrottrust.org Hayle, Cornwall Stillwater, MN 55082 TR27 4HB, United Kingdom Tel: 651 275 1877 Tel: (44) 1736 753365 Fax: 651 275 1891 Fax (44) 1736 751028 Island Press Box 7, Covelo, California 95428, USA Tel: 800 828 1302, 707 983 6432 Fax: 707 983 6414 E-mail: [email protected] Order on line: www.islandpress.org The views expressed in this publication are those of the authors and do not necessarily reflect those of IUCN or the Species Survival Commission. Published by: IUCN, Gland, Switzerland, and Cambridge, UK. Copyright: © 2000 International Union for Conservation of Nature and Natural Resources and the World Parrot Trust Reproduction of this publication for educational and other non-commercial purposes is authorised without prior written permission from the copyright holders provided the source is fully acknowledged. -
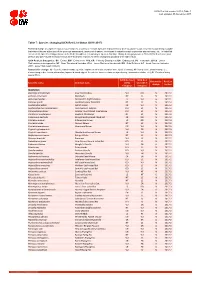
Table 7 Last Updated: 05 December 2017
IUCN Red List version 2017-3: Table 7 Last Updated: 05 December 2017 Table 7: Species changing IUCN Red List Status (2016-2017) Published listings of a species' status may change for a variety of reasons (genuine improvement or deterioration in status; new information being available that was not known at the time of the previous assessment; taxonomic changes; corrections to mistakes made in previous assessments, etc. To help Red List users interpret the changes between the Red List updates, a summary of species that have changed category between 2016 (IUCN Red List version 2016.3) and 2017 (IUCN Red List version 2017-3) and the reasons for these changes is provided in the table below. IUCN Red List Categories: EX - Extinct, EW - Extinct in the Wild, CR - Critically Endangered, EN - Endangered, VU - Vulnerable, LR/cd - Lower Risk/conservation dependent, NT - Near Threatened (includes LR/nt - Lower Risk/near threatened), DD - Data Deficient, LC - Least Concern (includes LR/lc - Lower Risk, least concern). Reasons for change: G - Genuine status change (genuine improvement or deterioration in the species' status); N - Non-genuine status change (i.e., status changes due to new information, improved knowledge of the criteria, incorrect data used previously, taxonomic revision, etc.); E - Previous listing was an Error. IUCN Red List IUCN Red Reason for Red List Scientific name Common name (2016) List (2017) change version Category Category MAMMALS Allactaga tetradactyla Four-toed Jerboa VU DD N 2017-2 Antilope cervicapra Blackbuck NT LC N 2017-2 -

WB3-Updateposted
Scientific Name Common Name Book Sex N Mean S.D. Min Max Season Location Source Comments Family Apterygidae Status Apteryx australis Southern Brown Kiwi E M 15 2120 1720 2730 New Zealand Colbourne & Kleinpaste 1983 F 31 2540 2060 3850 Apteryx australis lawryi Southern Brown Kiwi B M 9 2720 2300 3060 Stewart Island, New ZealandMarchant & Higgins 1990 F 10 3115 267 2700 3500 Apteryx rowi Okarito Brown Kiwi N M 49 1924 157 1575 2250 South Island, New Zealand Tennyson et al. 2003 F 51 2650 316 1950 3570 Apteryx mantelli North Island Brown Kiwi N F 4 2725 2060 3433 New Zealand Marchant & Higgins 1990 split in Clements Family Tinamidae Nothocercus julius Tawny-breasted Tinamou N F 1 693 Peru Specimens Mus. SWern Biology Nothocercus nigrocapillus Hooded Tinamou N M 1 540 Gomes and Kirwan 2014a Crypturellus brevirostris Rusty Tinamou N M 2 221 209 233 Guyana Robbins et al. 2007 F 1 295 Crypturellus casiquiare Barred Tinamou N F 1 250 Columbia Echeverry-Galvis, unpublished Rhynchotus maculicollis Huayco Tinamou N U 10 868 860 900 Fiora 1933a split by Clements 2008 Tinamotis ingoufi Patagonian Tinamou N U 900 1000 Gomes & Kirwan 2014b Family Spheniscidae Eudyptes moseleyi Tristan Penguin N B 11 2340 517 1400 2950 B Amsterdam Is., Indian OceanDrabek & Tremblay 2000 split from E. chrysocome by Clements 2012 Family Diomedeidae Diomedea antipodensis gibsoni Wandering Albatross N M 35 6800 5500 8600 Auckland Islands Tickell 2000 split by del Hoyo et al. 2013 F 36 5800 4600 7300 Diomedea antipodensis Wandering Albatross N M 10 7460 Antipodes Islands Tickell 2000 split by del Hoyo et al. -

French Polynesia, Pitcairn & Henderson
Tuamotu Sandpiper in the Tuamotus – the Holy Grail of South Pacific birding (Phil Tizzard) FRENCH POLYNESIA, PITCAIRN & HENDERSON 4/12 – 28/31 OCTOBER 2014 LEADER: DEREK SCOTT Birdquest’s third tour to remote French Polynesia, Pitcairn and Henderson was, like its predecessors, a great success. An eight-day pre-tour extension took us to five islands in the Marquesas; the main tour began with a day’s birding in Tahiti before travelling on to the Gambier Islands to join the R.V. Braveheart for a two-week cruise to the remote Pitcairn Islands and Actaeon Group of the Tuamotus; and the short post-tour extension gave us two days on the lovely little island of Rimatara in the Austral Islands. Thanks to the efficiency of Air Tahiti, French Polynesia’s domestic airline, our inter-island hopping all went very smoothly, and thanks to the skill and determination of Matt and his crew on the Braveheart, we were able to make landings on all the 1 BirdQuest Tour Report: French Polynesia, Pitcairn & Henderson www.birdquest-tours.com islands on our cruise itinerary, despite some unfavourable winds and choppy seas. By enlisting the help of local experts and guides for some of the trickier birds, we had no difficulty in finding all 24 of the endemic land birds on our list, while by putting in long hours of sea-watching from the deck of the Braveheart, we saw most of the seabirds that we were expecting plus a few that we weren’t. No fewer than 25 of the 68 naturally occurring species that we recorded are currently listed in Threatened Birds of the World as critically endangered, endangered or vulnerable, a sad reflection of the disastrous impact that human colonization and the associated introduction of rats has had on the native fauna of the eastern Polynesian islands. -

Spectrophotometric Studies of Avian Plumage Coloration with Special Emphasis on the Ultraviolet Range
View metadata, citation and similar papers at core.ac.uk brought to you by CORE Spectrophotometric Studies of Avian Plumage Coloration with Special Emphasis on the Ultraviolet Range Dissertation zur Erlangung des Doktorgrades (Dr. rer. nat.) der Mathematisch Naturwissenschaftlichen Fakultät der Rheinischen Friedrich-Wilhelms-Universität Bonn vorgelegt von Peter Mullen aus Furtwangen Bonn 2006 Angefertigt mit Genehmigung der Mathematisch-Naturwissenschaftlichen Fakultät der Rheinischen Friedrich-Wilhelms-Universität Bonn 1. Referent: Prof. Dr. K.-L.Schuchmann 2. Referent: Prof. Dr. H. Bleckmann Tag der Promotion: 08.02.2007 Diese Dissertation ist auf dem Hochschulschriftenserver der ULB Bonn http://hss.ulb.uni-bonn.de/diss_online elektronisch publiziert. Erscheinungsjahr: 2007 Contents 1. Studies on UV reflection in feathers of some 1000 bird species – are UV- peaks in feathers correlated with VS/UVS cones?...........................................10 Introduction ..................................................................................................................... 10 Methods ........................................................................................................................... 12 Results ............................................................................................................................. 14 Discussion........................................................................................................................ 22 Summary......................................................................................................................... -

Status and Conservation of the Vini Lorikeets of French Polynesia
STATUS AND CONSERVATION OF THE VINI LORIKEETS OF FRENCH POLYNESIA Report to the LORO PARQUE FOUNDATION & CONSERVATION DES ESPÈCES ET DES POPULATIONS ANIMALES (CEPA) JUNE 2006 MARC ZIEMBICKI PHILIPPE RAUST Vini lorikeets of French Polynesia 2 TABLE OF CONTENTS EXECUTIVE SUMMARY ......................................................................................................... 4 RÉSUMÉ EXÉCUTIF................................................................................................................. 5 ACKNOWLEDGEMENTS......................................................................................................... 6 ACRONYMS............................................................................................................................... 6 CITATION & CONTACT DETAILS......................................................................................... 6 INTRODUCTION ....................................................................................................................... 7 OBJECTIVES.............................................................................................................................. 8 PROJECT SITES......................................................................................................................... 9 Tuamotu archipelago ........................................................................................................... 9 Marquesas Islands...............................................................................................................