Summary of Responses
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-
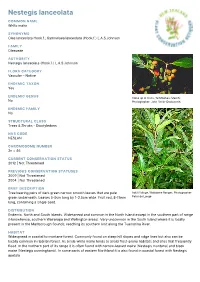
Nestegis Lanceolata
Nestegis lanceolata COMMON NAME White maire SYNONYMS Olea lanceolata Hook.f.; Gymnelaea lanceolata (Hook.f.) L.A.S.Johnson FAMILY Oleaceae AUTHORITY Nestegis lanceolata (Hook.f.) L.A.S.Johnson FLORA CATEGORY Vascular – Native ENDEMIC TAXON Yes ENDEMIC GENUS Close up of fruits, Te Moehau (March). No Photographer: John Smith-Dodsworth ENDEMIC FAMILY No STRUCTURAL CLASS Trees & Shrubs - Dicotyledons NVS CODE NESLAN CHROMOSOME NUMBER 2n = 46 CURRENT CONSERVATION STATUS 2012 | Not Threatened PREVIOUS CONSERVATION STATUSES 2009 | Not Threatened 2004 | Not Threatened BRIEF DESCRIPTION Tree bearing pairs of dark green narrow smooth leaves that are pale Adult foliage, Waitakere Ranges. Photographer: green underneath. Leaves 5-9cm long by 1-2.5cm wide. Fruit red, 8-11mm Peter de Lange long, containing a single seed. DISTRIBUTION Endemic. North and South Islands. Widespread and common in the North Island except in the southern part of range (Horowhenua, southern Wairarapa and Wellington areas). Very uncommon in the South Island where it is locally present in the Marlborough Sounds, reaching its southern limit along the Tuamarina River. HABITAT Widespread in coastal to montane forest. Commonly found on steep hill slopes and ridge lines but also can be locally common in riparian forest. As a rule white maire tends to avoid frost-prone habitats and sites that frequently flood. In the northern part of its range it is often found with narrow-leaved maire (Nestegis montana) and black maire (Nestegis cunninghamii). In some parts of eastern Northland it is also found in coastal forest with Nestegis apetala. FEATURES Stout gynodioecious spreading tree up to 20 m tall usually forming a domed canopy; trunk up to c. -

Torus-Bearing Pit Membranes in Species of Osmanthus
IAWA Journal, Vol. 31 (2), 2010: 217–226 TORUS-BEARING PIT MEMBRANES IN SPECIES OF OSMANTHUS Roland Dute1, 5, David Rabaey2, John Allison1 and Steven Jansen3, 4 SUMMARY Torus thickenings of pit membranes are found not only in gymnosperms, but also in certain genera of dicotyledons. One such genus is Osmanthus. Wood from 17 species of Osmanthus was searched for tori. Fourteen spe- cies from three of the four sections investigated possessed these thick- enings. Ten of the species represent new records. Only the three New Caledonian species of Section Notosmanthus lacked tori. This observation in combination with other factors serves to isolate this section from the remainder of the genus. Key words: Notelaea, Osmanthus, pit membrane, torus. INTRODUCTION Osmanthus is a member of the Oleaceae (Olive Family), a moderate-sized family with circa 24 genera and 615 spp. (Stevens 2001). There are 33 natural species within the genus (Table 1; Xiang et al. 2008), and they show a tropical to temperate distribution (Bailey & Bailey 1976; Denk et al. 2001; Xiang et al. 2008). Generally, individual shrubs are found in moist forests (Hsieh et al. 1998; Chou et al. 2000; Denk et al. 2001), frequently on steep slopes (Green 1958). In some instances populations are sub- ject to monsoon conditions and hence to periods of relatively low rainfall (Yang et al. 2008; Hua 2008). Tori are thickenings of intervascular pit membranes found not only in gymnosperm, but also in angiosperm wood, including that of Osmanthus (Dute et al. 2008; Dute et al. 2010). Tori were first observed in O. fragrans, O. -

Patterns of Flammability Across the Vascular Plant Phylogeny, with Special Emphasis on the Genus Dracophyllum
Lincoln University Digital Thesis Copyright Statement The digital copy of this thesis is protected by the Copyright Act 1994 (New Zealand). This thesis may be consulted by you, provided you comply with the provisions of the Act and the following conditions of use: you will use the copy only for the purposes of research or private study you will recognise the author's right to be identified as the author of the thesis and due acknowledgement will be made to the author where appropriate you will obtain the author's permission before publishing any material from the thesis. Patterns of flammability across the vascular plant phylogeny, with special emphasis on the genus Dracophyllum A thesis submitted in partial fulfilment of the requirements for the Degree of Doctor of philosophy at Lincoln University by Xinglei Cui Lincoln University 2020 Abstract of a thesis submitted in partial fulfilment of the requirements for the Degree of Doctor of philosophy. Abstract Patterns of flammability across the vascular plant phylogeny, with special emphasis on the genus Dracophyllum by Xinglei Cui Fire has been part of the environment for the entire history of terrestrial plants and is a common disturbance agent in many ecosystems across the world. Fire has a significant role in influencing the structure, pattern and function of many ecosystems. Plant flammability, which is the ability of a plant to burn and sustain a flame, is an important driver of fire in terrestrial ecosystems and thus has a fundamental role in ecosystem dynamics and species evolution. However, the factors that have influenced the evolution of flammability remain unclear. -

Breeding Systems and Reproduction of Indigenous Shrubs in Fragmented
Copyright is owned by the Author of the thesis. Permission is given for a copy to be downloaded by an individual for the purpose of research and private study only. The thesis may not be reproduced elsewhere without the permission of the Author. Breeding systems and reproduction of indigenous shrubs in fragmented ecosystems A thesis submitted in partial fulfilment of the requirements for the degree of Doctor of Philosophy III Plant Ecology at Massey University by Merilyn F Merrett .. � ... : -- �. � Massey University Palrnerston North, New Zealand 2006 Abstract Sixteen native shrub species with various breeding systems and pollination syndromes were investigated in geographically separated populations to determine breeding systems, reproductive success, population structure, and habitat characteristics. Of the sixteen species, seven are hermaphroditic, seven dioecious, and two gynodioecious. Two of the dioecious species are cryptically dioecious, producing what appear to be perfect, hermaphroditic flowers,but that functionas either male or female. One of the study species, Raukauaanomalus, was thought to be dioecious, but proved to be hermaphroditic. Teucridium parvifolium, was thought to be hermaphroditic, but some populations are gynodioecious. There was variation in self-compatibility among the fo ur AIseuosmia species; two are self-compatible and two are self-incompatible. Self incompatibility was consistent amongst individuals only in A. quercifolia at both study sites, whereas individuals in A. macrophylia ranged from highly self-incompatible to self-compatible amongst fo ur study sites. The remainder of the hermaphroditic study species are self-compatible. Five of the species appear to have dual pollination syndromes, e.g., bird-moth, wind-insect, wind-animal. High levels of pollen limitation were identified in three species at fo ur of the 34 study sites. -

9:00 Am PLACE
CARTY S. CHANG INTERIM CHAIRPERSON DAVID Y. IGE BOARD OF LAND AND NATURAL RESOURCES GOVERNOR OF HAWAII COMMISSION ON WATER RESOURCE MANAGEMENT KEKOA KALUHIWA FIRST DEPUTY W. ROY HARDY ACTING DEPUTY DIRECTOR – WATER AQUATIC RESOURCES BOATING AND OCEAN RECREATION BUREAU OF CONVEYANCES COMMISSION ON WATER RESOURCE MANAGEMENT STATE OF HAWAII CONSERVATION AND COASTAL LANDS CONSERVATION AND RESOURCES ENFORCEMENT DEPARTMENT OF LAND AND NATURAL RESOURCES ENGINEERING FORESTRY AND WILDLIFE HISTORIC PRESERVATION POST OFFICE BOX 621 KAHOOLAWE ISLAND RESERVE COMMISSION LAND HONOLULU, HAWAII 96809 STATE PARKS NATURAL AREA RESERVES SYSTEM COMMISSION MEETING DATE: April 27, 2015 TIME: 9:00 a.m. PLACE: Department of Land and Natural Resources Boardroom, Kalanimoku Building, 1151 Punchbowl Street, Room 132, Honolulu. AGENDA ITEM 1. Call to order, introductions, move-ups. ITEM 2. Approval of the Minutes of the June 9, 2014 N atural Area Reserves System Commission Meeting. ITEM 3. Natural Area Partnership Program (NAPP). ITEM 3.a. Recommendation to the Board of Land and Natural Resources approval for authorization of funding for The Nature Conservancy of Hawaii for $663,600 during FY 16-21 for continued enrollment in the natural area partnership program and acceptance and approval of the Kapunakea Preserve Long Range Management Plan, TMK 4-4-7:01, 4-4-7:03, Lahaina, Maui. ITEM 3.b. Recommendation to the Board of Land and Natural Resources approval for authorization of funding for The Nature Conservancy of Hawaii for $470,802 during FY 16-21 for continued enrollment in the natural area partnership program and acceptance and approval of the Pelekunu Long Range Management Plan, TMK 5-4- 3:32, 5-9-6:11, Molokai. -
To Obtain Approval to Release New Organisms (Through Importing for Release Or Releasing from Containment)
APPLICATION FORM Release To obtain approval to release new organisms (Through importing for release or releasing from containment) Send to Environmental Protection Authority preferably by email ([email protected]) or alternatively by post (Private Bag 63002, Wellington 6140) Payment must accompany final application; see our fees and charges schedule for details. Figure 1. Tree privet displacing coastal pohutukawa forest. Application Number APP202262 Date 13 January 2015 www.epa.govt.nz 2 Application Form Approval to release a new organism Completing this application form 1. This form has been approved under section 34 of the Hazardous Substances and New Organisms (HSNO) Act 1996. It covers the release without controls of any new organism (including genetically modified organisms (GMOs)) that is to be imported for release or released from containment. It also covers the release with or without controls of low risk new organisms (qualifying organisms) in human and veterinary medicines. If you wish to make an application for another type of approval or for another use (such as an emergency, special emergency, conditional release or containment), a different form will have to be used. All forms are available on our website. 2. It is recommended that you contact an Advisor at the Environmental Protection Authority (EPA) as early in the application process as possible. An Advisor can assist you with any questions you have during the preparation of your application including providing advice on any consultation requirements. 3. Unless otherwise indicated, all sections of this form must be completed for the application to be formally received and assessed. If a section is not relevant to your application, please provide a comprehensive explanation why this does not apply. -

Lamiales – Synoptical Classification Vers
Lamiales – Synoptical classification vers. 2.6.2 (in prog.) Updated: 12 April, 2016 A Synoptical Classification of the Lamiales Version 2.6.2 (This is a working document) Compiled by Richard Olmstead With the help of: D. Albach, P. Beardsley, D. Bedigian, B. Bremer, P. Cantino, J. Chau, J. L. Clark, B. Drew, P. Garnock- Jones, S. Grose (Heydler), R. Harley, H.-D. Ihlenfeldt, B. Li, L. Lohmann, S. Mathews, L. McDade, K. Müller, E. Norman, N. O’Leary, B. Oxelman, J. Reveal, R. Scotland, J. Smith, D. Tank, E. Tripp, S. Wagstaff, E. Wallander, A. Weber, A. Wolfe, A. Wortley, N. Young, M. Zjhra, and many others [estimated 25 families, 1041 genera, and ca. 21,878 species in Lamiales] The goal of this project is to produce a working infraordinal classification of the Lamiales to genus with information on distribution and species richness. All recognized taxa will be clades; adherence to Linnaean ranks is optional. Synonymy is very incomplete (comprehensive synonymy is not a goal of the project, but could be incorporated). Although I anticipate producing a publishable version of this classification at a future date, my near- term goal is to produce a web-accessible version, which will be available to the public and which will be updated regularly through input from systematists familiar with taxa within the Lamiales. For further information on the project and to provide information for future versions, please contact R. Olmstead via email at [email protected], or by regular mail at: Department of Biology, Box 355325, University of Washington, Seattle WA 98195, USA. -

Flowering Plants
Thonner’s analytical key to the families of flowering plants R. Geesink A.J.M. Leeuwenberg C.E. Ridsdale J.F. Veldkamp PUDOC, Centre for Agricultural Leiden University Press Publishing and Documentation The Hague/Boston/London, Wageningen, 1981 1981 R. Geesink-Rijksherbarium, Leiden, Netherlands A. J. M. - Laboratorium Planten- Leeuwenberg voor Plantensystematiek en geografie, Agricultural University, Wageningen, Netherlands C. E. Ridsdale-B. A. Krukoff Botanist of Malesian Botany, Rijksherbarium, Leiden, Netherlands J. F. Veldkamp-Rijksherbarium, Leiden, Netherlands This volume is listed in the Library of Congress Cataloging in Publication Data This is a translated and revised edition of: Anleitung zum Bestimmen der Familien der Bliitenpflanzen, 2nd. ed. 1917, Friedlander, Berlin ISBN 90-6021-479-X ISBN 90 - 6021-462- 5 (series Leiden University Press) © Centre for agricultural publishing and documentation, PUDOC, Wageningen 1981 and Martinus Nijhoff Publishers, The Hague, 1981 stored All rights reserved. No part of this publication may be reproduced, in a retrieval system, or transmitted in any form or by any means, mechanical, photocopying, recording, or otherwise, without the prior written permission of the publishers, Martinus Nijhoff Publishers, P.O. Box 566, 2501 CN The Hague, The Netherlands, and PUDOC, P.O. Box 4, 6700 AA Wageningen, The Netherlands Printed in the Netherlands Contents Preface to the2nd edition (1917) vii Introduction viii Acknowledgements x Franz Thonner-Life (1863-1928) xii Franz Thonner-Bibliography xv Franz Thonner-Derivedworks xviii Franz Thonner-Eponymy xx The Key - Introduction and Notes xxii Scheme for a diagnostic description xxvi Concise key to the major groupings 1 Key to the families 3 Glossary 198 Index 214 'All plants are hybrids, but some are greater bastards than others' Franz Thonner. -

NZ BOT SOC No 111 March 2013
NEW ZEALAND BOTANICAL SOCIETY NEWSLETTER NUMBER 111 March 2013 New Zealand Botanical Society President: Anthony Wright Secretary/Treasurer: Ewen Cameron Committee: Bruce Clarkson, Colin Webb, Carol West Address: c/- Canterbury Museum Rolleston Avenue CHRISTCHURCH 8013 URL: www.nzbotanicalsociety.org.nz Subscriptions The 2013 ordinary and institutional subscriptions are $25 (reduced to $18 if paid by the due date on the subscription invoice). The 2012 student subscription, available to full-time students, is $12 (reduced to $9 if paid by the due date on the subscription invoice). Back issues of the Newsletter are available at $7.00 each. Since 1986 the Newsletter has appeared quarterly in March, June, September and December. New subscriptions are always welcome and these, together with back issue orders, should be sent to the Secretary/Treasurer (address above). Subscriptions are due by 28 February each year for that calendar year. Existing subscribers are sent an invoice with the December Newsletter for the next years subscription which offers a reduction if this is paid by the due date. If you are in arrears with your subscription a reminder notice comes attached to each issue of the Newsletter. Deadline for next issue The deadline for the June 2013 issue is 25 May 2013. Please post contributions to: Lara Shepherd Museum of New Zealand Te Papa Tongarewa 169 Tory St Wellington 6021 Send email contributions to [email protected]. Files are preferably in MS Word, as an open text document (Open Office document with suffix “.odt”) or saved as RTF or ASCII. Macintosh files can also be accepted. Graphics can be sent as TIF JPG, or BMP files; please do not embed images into documents. -

Delissea Subcordata
Plants ‘Oha Delissea subcordata SPECIES STATUS: Federally Listed as Endangered Genetic Safety Net Species John K. Obata © Smithsonian Inst., 2005 Hawai‘i Natural Heritage Ranking ‐ Critically Imperiled (G1) Endemism – O‘ahu Critical Habitat ‐ Designated SPECIES INFORMATION: Delissea subcordata, a member of the bellflower family (Campanulaceae), is a branched or unbranched shrub 1 ‐ 3 m tall. Leaves ovate or ovate‐ lanceolate, blades 12 ‐ 30 cm long, 6 ‐ 17 cm wide, margins crenate, denticulate, serrate, or doubly serrate, the teeth incurved, or occasionally irregularly laciniate with 1 ‐ 6 triangular lobes 10 ‐ 18 mm long toward the base, apex obtuse, acute, or acuminate, base cordate, subcordate, or truncate, petioles 6 ‐ 18 cm long. Inflorescences 6 ‐ 18 flowered, glabrous, peduncles 40 ‐ 100 mm long, pedicels 13 ‐ 18 mm long; hypanthium ovoid to cylindrical; calyx lobes subulate, 0.5 ‐ 1 mm long; corolla white or greenish white, curved, 45 ‐ 60 mm long, with 1 dorsal knob; anthers glabrous. Berries ovoid, 12 ‐16 mm long, 8 ‐ 12 mm in diameter. Seeds ca. 1 mm long. Occurring in diverse mesic forest, 250 ‐ 400 m, Ko‘olau Mountains, and 450 ‐ 550 m, Wai‘anae Mountains, O‘ahu. DISTRIBUTION: The persistence of this plant in the Wai‘anae Mountains is well documented. However, it has not been collected in the Ko‘olau Mountains since 1934 and may be extinct there. ABUNDANCE: Currently 60‐70 plants observed. Delissea subcordata is now known only from the Wai‘anae Mountains in nine populations distributed from Kawaiu Gulch in the Kealia land section in the northern Wai‘anae Mountains to the north branch of North Palawai Gulch about 20 km (12 mi) to the south. -

Plants Delissea Rhytidosperma
Plants Delissea rhytidosperma SPECIES STATUS: Federally Listed as Endangered Genetic Safety Net Species W.L. Wagner © Smithsonian Inst., 2005 Hawai‘i Natural Heritage Ranking‐ Critically Imperiled (G1) Endemism – Kaua‘i Critical Habitat ‐ Designated SPECIES INFORMATION: Delissea rhytidosperma, a member of the bellflower family, is a branched shrub 1.6 to 8.2 ft (0.5 to 2‐5 m) tall. The lance‐shaped or elliptic leaves are 3.1 to 7.5 in (8 to 19 cm) long and 0.8 to 2.2 in (2 to 5.5 cm) wide and have toothed margins. Clusters of 5 to 12 flowers are borne on stalks 0.4 to 0.8 in (1 to 2 cm) long; each flower has a stalk 0.3 to 0.5 in (8 to 13 mm) long. The greenish white (sometimes pale purple) corolla is 0.6 to 0.8 in (14 to 20 mm) long. The stamens are hairless, except for a small patch of hair at the base of the anthers. The nearly spherical dark purple fruits are 0.3 to 0.5 in (7 to 12 mm) long and contain numerous white seeds. This species differs from other taxa of the genus by the shape, length, and margins of the leaves and by having hairs at the base of the anthers. DISTRIBUTION: Historically, Delissea rhvtidosperma was known from scattered locations throughout the island of Kaua‘i. Populations ranged as far north as Wainiha and Limahuli Valleys, as far east as Kapaa and Kealia, and as far south as Haupu Range between the elevations of 1,000 and 3,000 ft (300 and 1,000 m). -

A Synoptical Classification of the Lamiales
Lamiales – Synoptical classification vers. 2.0 (in prog.) Updated: 13 December, 2005 A Synoptical Classification of the Lamiales Version 2.0 (in progress) Compiled by Richard Olmstead With the help of: D. Albach, B. Bremer, P. Cantino, C. dePamphilis, P. Garnock-Jones, R. Harley, L. McDade, E. Norman, B. Oxelman, J. Reveal, R. Scotland, J. Smith, E. Wallander, A. Weber, A. Wolfe, N. Young, M. Zjhra, and others [estimated # species in Lamiales = 22,000] The goal of this project is to produce a working infraordinal classification of the Lamiales to genus with information on distribution and species richness. All recognized taxa will be clades; adherence to Linnaean ranks is optional. Synonymy is very incomplete (comprehensive synonymy is not a goal of the project, but could be incorporated). Although I anticipate producing a publishable version of this classification at a future date, my near-term goal is to produce a web-accessible version, which will be available to the public and which will be updated regularly through input from systematists familiar with taxa within the Lamiales. For further information on the project and to provide information for future versions, please contact R. Olmstead via email at [email protected], or by regular mail at: Department of Biology, Box 355325, University of Washington, Seattle WA 98195, USA. Lamiales – Synoptical classification vers. 2.0 (in prog.) Updated: 13 December, 2005 Acanthaceae (~201/3510) Durande, Notions Elém. Bot.: 265. 1782, nom. cons. – Synopsis compiled by R. Scotland & K. Vollesen (Kew Bull. 55: 513-589. 2000); probably should include Avicenniaceae. Nelsonioideae (7/ ) Lindl. ex Pfeiff., Nomencl.