Using Beef Cattle Genetics to Manage
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Postweaning Growth and Carcass Traits in Crossbred Cattle from Hereford, Angus, Brangus, Beefmaster, Bonsmara, and Romosinuano Maternal Grandsires E
University of Nebraska - Lincoln DigitalCommons@University of Nebraska - Lincoln Roman L. Hruska U.S. Meat Animal Research U.S. Department of Agriculture: Agricultural Center Research Service, Lincoln, Nebraska 2010 Postweaning growth and carcass traits in crossbred cattle from Hereford, Angus, Brangus, Beefmaster, Bonsmara, and Romosinuano maternal grandsires E. Casas USDA, ARS, US Meat Animal Research Center, [email protected] R. M. Thallman USDA-ARS Meat Animal Research Center, [email protected] L. A. Kuehn USDA- ARS, US Meat Animal Research Center, [email protected] L. V. Cundiff US Meat Animal Research Center, [email protected] Follow this and additional works at: http://digitalcommons.unl.edu/hruskareports Casas, E.; Thallman, R. M.; Kuehn, L. A.; and Cundiff, L. V., "Postweaning growth and carcass traits in crossbred cattle from Hereford, Angus, Brangus, Beefmaster, Bonsmara, and Romosinuano maternal grandsires" (2010). Roman L. Hruska U.S. Meat Animal Research Center. 374. http://digitalcommons.unl.edu/hruskareports/374 This Article is brought to you for free and open access by the U.S. Department of Agriculture: Agricultural Research Service, Lincoln, Nebraska at DigitalCommons@University of Nebraska - Lincoln. It has been accepted for inclusion in Roman L. Hruska U.S. Meat Animal Research Center by an authorized administrator of DigitalCommons@University of Nebraska - Lincoln. Published December 4, 2014 Postweaning growth and carcass traits in crossbred cattle from Hereford, Angus, Brangus, Beefmaster, Bonsmara, and Romosinuano maternal grandsires1,2 E. Casas,3 R. M. Thallman, L. A. Kuehn, and L. V. Cundiff USDA, ARS, US Meat Animal Research Center, Clay Center, NE 68933 ABSTRACT: The objective of this study was to char- est marbling scores when compared with other grand- acterize breeds representing diverse biological types for sire breeds. -
Proc1-Beginning Chapters.Pmd
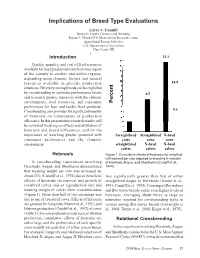
Implications of Breed Type Evaluations Larry V. Cundiff Research Leader, Genetics and Breeding Roman L. Hruska U.S. Meat Animal Research Center Agricultural Research Service U.S. Department of Agriculture Clay Center, NE ()(,) Introduction 23.3 Quality, quantity, and cost of feed resources available for beef production vary from one region of the country to another and within regions, depending upon climatic factors and natural 23.3 resources available in specific production 14.8 situations. Diversity among breeds can be exploited by crossbreeding to optimize performance levels 8.5 and to match genetic resources with the climatic environment, feed resources, and consumer Percent preferences for lean and tender beef products. 8.5 Crossbreeding also provides for significant benefits 14.8 of heterosis on components of production efficiency. In this presentation, research results will be reviewed focusing on effects and utilization of 8.5 8.5 heterosis and breed differences, and on the importance of matching genetic potential with Straightbred Straightbred X-bred consumer preferences and the climatic cows cows cows environment. straightbred X-bred X-bred calves calves calves Heterosis Figure 1. Cumulative effects of heterosis for weight of calf weaned per cow exposed to breeding in crosses A crossbreeding experiment involving of Hereford, Angus, and Shorthorns (Cundiff et al., Herefords, Angus, and Shorthorns demonstrated 1974). that weaning weight per cow was increased by about 23% (Cundiff et al., 1974) due to beneficial was significantly greater than that of either effects of heterosis on survival and growth of straightbred Angus or Herefords (Nunez et al., crossbred calves and on reproduction rate and 1991; Cundiff et al., 1992). -

For Immediate Release Brangus Are Not “Eared” Cattle
For Immediate Release Contact: Doc or Patricia Spitzer FAIR PLAY, SC [email protected] or November 8, 2011 (864)972-9140 or (864)710-0257 Brangus Are Not “Eared” Cattle First and foremost we need to wrap our minds around the fact that God created cattle, he did not create breeds. And while in some cases natural barriers such as oceans and mountain ranges did affect genetic selection, for the most part it is humans who created breeds. And, if you go back far enough in history there really are no pure breeds, only our inflated misconceptions that they exist. That being said, there are two species that make up all cattle of the world; Bos Taurus cattle are primarily cattle populations that originated in the more temperate climates and Bos Indicus cattle populations developed in the more tropical regions of the world. Generally the US beef industry further subdivides Bos Taurus beef cattle into two groups. Continental Breeds of cattle are those breeds originating on the European Continent while British Breeds were originally from selections of bovine populations from the British Isle. It is also typical of US producers to wrongly lump all Bos Indicus breeds of cattle together. This is rather astounding as there are more recognized different breeds of Bos Indicus derivation scattered around the world than specific breeds of Bos Taurus derivation. Americans have also further compounded the confusion by creating new breeds by crossing a variety of specifically recognized Bos Indicus cattle to create the American Brahman and crossing cattle of Bos Indicus and Bos Taurus origin to create what some refer to as the American Breeds. -

Purebred Livestock Registry Associations
Purebred livestock registry associations W. Dennis Lamm1 COLORADO STATE UNIVERSITY EXTENSION SERVICE no. 1.217 Beef Devon. Devon Cattle Assn., Inc., P.O. Box 628, Uvalde, TX 78801. Mrs. Cammille Hoyt, Sec. Phone: American. American Breed Assn., Inc., 306 512-278-2201. South Ave. A, Portales, NM 88130. Mrs. Jewell Dexter. American Dexter Cattle Assn., P.O. Jones, Sec. Phone: 505-356-8019. Box 56, Decorah, IA 52l01. Mrs. Daisy Moore, Amerifax. Amerifax Cattle Assn., Box 149, Exec. Sec. Phone: 319-736-5772, Hastings, NE 68901. John Quirk, Pres. Phone Friesian. Beef Friesian Society, 213 Livestock 402-463-5289. Exchange Bldg., Denver, CO 80216. Maurice W. Angus. American Angus Assn., 3201 Freder- Boney, Adm. Dir. Phone: 303-587-2252. ick Blvd., St. Joseph, MO 64501. Richard Spader, Galloway. American Galloway Breeders Assn., Exec. Vice. Pres. Phone: 816-233-3101. 302 Livestock Exchange Bldg., Denver, CO 80216. Ankina. Ankina Breeders, Inc., 5803 Oaks Rd,. Cecil Harmon, Pres. Phone: 303-534-0853. Clayton, OH 45315. James K. Davis, Ph.D., Pres. Galloway. Galloway Cattle Society of Amer- Phone: 513-837-4128. ica, RFD 1, Springville, IA 52336. Phone: 319- Barzona. Barzona Breeders Assn. of America, 854-7062. P.O. Box 631, Prescott, AZ 86320. Karen Halford, Gelbvieh. American Gelbvieh Assn., 5001 Na- Sec. Phone: 602-445-2290. tional Western Dr., Denver, CO 80218. Daryl W. Beefalo. American Beefalo Breeders, 1661 E. Loeppke, Exec. Dir. Phone: 303-296-9257. Brown Rd., Mayville 22, MI 48744. Phone: 517-843- Hays Convertor. Canadian Hays Convertor 6811. Assn., 6707 Elbow Dr. SW, Suite 509, Calgary, Beefmaster. -

Breeding Heifer Validation Program Major Livestock Show Breed Eligibility Guide
Breeding Heifer Validation Program Major Livestock Show Breed Eligibility Guide This document is only to serve as a resource for determining the eligibility of specific breeds at major livestock shows. This is not a complete list of breeds eligible and livestock shows reserve the right to add/remove breeds at the discretion of show management. *Always refer to respective livestock show’s official rules and information. ü 1 ORB – Other Registered Breeds (Bos Taurus type, non-American), ARB – American Registered Breeds (Bos indicus type) ü 2 ORH – Other Recorded Heifer (Bos Taurus type, non-American) ü ARB – American Registered Breeds (Bos indicus type) ü n/a – means not applicable (i.e., breed not eligible for exhibition at show) BREED HLSR 1 SALE 1 SAN ANGELO 1 RODEO AUSTIN 1 FORT WORTH 2 STATE FAIR 1 HOT FAIR Angus Angus Angus Angus Angus Angus Angus Angus Beefmaster Beefmaster Beefmaster Beefmaster Beefmaster Beefmaster Beefmaster Beefmaster Beefmaster ARB ARB ARB ARB n/a ARB ARB E6 Certified (min 50% Beefmaster) Beefmaster ARB ARB ARB ARB n/a ARB ARB Advancer Braford ARB ARB ARB ARB n/a ARB ARB Braford F-1 ARB n/a ARB ARB n/a ARB ARB Brahman Grey Brahman OR Brahman ARB Brahman Brahman Grey Brahman OR Grey Brahman Red Brahman (Red & Grey (Red & Grey (Red & Grey Red Brahman OR Combined) Combined) Combined) Red Brahman Page 1 of 5 Breeding Heifer Validation Program Major Livestock Show Breed Eligibility Guide BREED HLSR 1 SALE 1 SAN ANGELO 1 RODEO AUSTIN 1 FORT WORTH 2 STATE FAIR 1 HOT FAIR Brahman ARB ARB ARB ARB n/a ARB ARB Golden Certified -

Impacts of Beef Cattle Shortcourses
Impacts of Beef Cattle Shortcourses Dr. Tim Olson Department of Animal Sciences University of Florida, Gainesville The first Breeder’s and Herdsmen’s Short - probably not such a good idea for the showman Course, the predecessor of the Beef Cattle Short either. Courses that have now been held for 50 years, was held on June 18th and 19th, 1951 at the University Mr. Joe Keefauver, an Angus breeder from of Florida. As the title of this first short course Tennessee, spoke on breeding and commented implies, the emphasis was on the production of that although the best cattle come from purebred cattle and the event was sponsored by linebreeding, most people can’t successfully do it the Florida Angus Breeders’ Association. About and most cattle aren’t good enough to be linebred 75 attended this first short course whose topics without running the risk of bringing out bad included fundamentals of feeding beef cattle by characteristics. A lot of emphasis was placed on Dr. R. S. Glasscock of the UF Animal Husbandry visual evaluation by the speakers, as might have Department, selection of breeding females by Joe been expected for the time, but Mr. Keefauver Keefauver, an Angus breeder from Tennessee and also added “You can’t tell for sure by looking, selection of a herd bull by Rae Ferrell, an Angus production is the ultimate test.” Mr. Keefauver breeder from Georgia. A local Gainesville was ahead of his time. Another speaker, however, veterinarian, Dr. Karl Owens discussed herd made the comment that “You won’t find a bull health issues including Bangs, Blackleg, Scours that is too short-legged.” Unfortunately this and Footrot. -

A Compilation of Research Results Involving Tropically Adapted Beef Cattle Breeds
A COMPILATION OF RESEARCH RESULTS INVOLVING TROPICALLY ADAPTED BEEF CATTLE BREEDS S-243 and S-277 Multistate Research Projects Southern Cooperative Series Bulletin 405 http://www.lsuagcenter.com/en/crops_livestock/livestock/beef_cattle/breeding_genetics/trpoical+breeds.htm Contact information: Dr. David G. Morrison, Associate Director Louisiana Agricultural Experiment Station P. O. Box 25055 Baton Rouge, Louisiana 70894-5055 Phone: 225-578-4182 FAX: 225-578-6032 Email: [email protected] ISBN: 1-58161-405-5 State Agricultural Experiment Stations do not discriminate on the basis of race, sex, color, religion, national origin, age, disability, or veteran status in provision of educational opportunities or employment opportunities and benefits. - 1 - Preface The Southern region of the U.S. contains approximately 42% of the nation’s beef cows and nearly 50% of its cow-calf producers. The region’s environment generally can be characterized as subtropical, i.e. hot, humid summers with ample rainfall supporting good forage production. Efficient cow-calf production in the humid South is dependent on heat and parasite tolerance and good forage utilization ability. Brahman and Brahman-derivative breeds generally possess these characteristics and excel in maternal traits. Consequently, they have been used extensively throughout the Southern Region in crossbreeding systems with Bos taurus breeds in order to exploit both breed complementarity and heterosis effects. However, several characteristics of Brahman and Brahman crossbred cattle, such as poor feedlot performance, lower carcass quality including meat tenderness, and poor temperament, whether real or perceived can result in economic discounts of these cattle. Therefore, determining genetic variation for economically important traits among Brahman and Brahman-derivative breeds and identifying tropically adapted breeds of cattle from other countries that may excel in their performance of economically important traits in Southern U.S. -

Florida Agriculture Statistical Directory
Dear Friends of Agriculture, It is my pleasure to present the 2008 Florida Agriculture Statistical Directory. This report presents a wealth of information about Florida’s vast and varied agricultural production through data that details land use, crop yields, commodity prices, crop rankings and more. This yearly report is invaluable to anyone who is involved in this dynamic business or who wants to better understand its complexities. The tables, charts and statistics contained in this report do an exceptional job of measuring the inputs and outputs, and presenting Florida agriculture in the context of “hard numbers.” But there is more to our state’s agricultural industry: our hard-working farmers, whose dedication, hard work and perseverance have made Florida agriculture into the diverse and highly productive industry that is respected throughout the globe. As evidenced by the ever-growing popularity of the “Fresh from Florida” label, consumers worldwide appreciate and seek out the quality products that our farmers provide. Maintaining these standards of excellence seldom comes easily as each year presents new challenges for Florida’s 40,000 commercial farmers. But, whether confronted by hurricanes, freezes, pests, diseases or fierce international competition, our state’s producers continually show that they are up to the test. Enterprising spirit, love of the land, and pride in their products are all hallmarks of the well- earned reputation of Florida’s farmers. In addition to enjoying the quality products that our farmers produce, Florida’s agricultural production benefits our state’s residents in other important ways as well. Florida agriculture has an overall economic impact estimated at more than $100 billion annually, making it a sound pillar of the state’s economy. -

History of Ibba
HISTORY OF IBBA How It All Began A review of the development of the Brangus breed would take us back beyond the founding of the American Brangus Breeders Association in 1949; however, registered Brangus descend from the foundation animals recorded that year or registered Brahman and Angus cattle enrolled since then. Much of the early work in crossing Angus and Brahman cattle was done at the USDA Experiment Station at Jeanerette, Louisiana -- the first crosses being made as far back as 1912. During the same period, Clear Creek Ranch of Welch, Oklahoma, the Essar Ranch of San Antonio, Texas, and a few individual breeders in other parts of the United States and Canada were also carrying on private experimental breeding programs. They were looking for a desirable beef-type animal that would retain the Brahman's natural ability to thrive under adverse conditions in combination with the excellent qualities for which the Angus was noted. The early breeders from 16 states and Canada met in Vinita, Oklahoma on July 19, 1949, and organized the American Brangus Breeders Association, later renamed the International Brangus Breeders Association, with headquarters in Kansas City, Missouri. One day following the formation of the American Brangus Breeders Association, the July 30, 1949 edition of the Vinita Daily Journal, Vinita, Oklahoma, published a front page article entitled "Pope is named president of Brangus Group." The article went on to read: "Other officers elected here as cattle organization is set up. Raymond Pope, co-owner and manager of Clear Creek Ranch in Northern Craig County was elected president of the American Brangus Breeders Association at the organization meeting for the group held Friday afternoon in Vinita. -

Development Simbrah Breed
Development of the Simbrah Breed What began as little more than an idea in the late The basic requirements for Simbrah cattle 1960’s on the part of a few cattlemen has now emerged are as follows: as a viable beef breed in the United States. The concept 1. Every Simbrah animal must contain of combining two of the most populous breeds of cattle a. minimum of 1/8 Simmental in the world, Brahman, which ranks number one in the b. minimum of 1/8 Brahman numbers, and Simmental, which ranks number two, c. maximum of 3/8 combination of other breeds resulted in the development of a breed known as Simbrah. Although a number of American beef breeds have been Examples of acceptable combinations could range developed using Brahman blood including the Santa from: 3/8 SM-1/4 BR-3/8 (maximum portion of other Gertrudis, Brangus and Beefmaster, the success of the breeds allowed) to 3/4 SM 1/8 BR (maximum portion Simbrah breed has been startling. of Simmental allowed) to 1/8 SM - 7/8 BM (min. Simmental allowed). Using two distinctly different breeds of cattle utilizes 2. The following criteria must be met to qualify the strengths of both breeds and because of the genetically as a purebred Simbrah: divergent background, maximizes hybrid vigor. Combining a. Simmental = 5/8 the strengths of the Brahman breed, which include b. Brahman = 3/8 longevity, heat tolerance, disease and insect resistance, c. Both parents must be ASA registered membership durability, grazing ability and calving ease with excellent of the American Simmental Association in 1977, traits from the Simmental, such as fertility, milking ability, with breeders registering 700 animals the first year rapid growth and early sexual maturity gives an excellent of existence. -

American Registered Breeds (Arb) Other Registered
AMERICAN REGISTERED BREEDS (ARB) 1. Open to heifers of Bos indicus type that have been registered or issued a certificate of recordation with a recognized breed association. The breeds that will be recognized are defined below: • Beefmaster Advancer - Animals of fifty percent (50%) or more Registered Beefmaster breeding and fifty percent (50%) or less of other Registered and DNA genotyped non-Beefmaster Beef cattle breeding. • Braford - Purebred, Heifers must be classified as Braford to be eligible to show. No F-1, multiple generation half blood, ¾ Hereford, ¾ Brahman, Single Bar or other percentage cattle will be accepted. • Brangus Premium Gold – Progeny of Registered Brangus or Red Brangus and any commercial animal. Must maintain a minimum of 50% registered Brangus/Red Brangus genetics. • Brangus Optimizer – Progeny resulting from mating Registered Brangus or Red Brangus and a registered animal from another beef breed recognized by the U.S. Beef Breeds Council, other than Angus or Red Angus. Must maintain a minimum of 50% registered Brangus genetics. • Brangus UltraBlack – Progeny of Registered Brangus and an Enrolled Angus • Brangus UltraRed- Progeny of Registered Red Brangus and an Enrolled Angus/Red Angus • Certified Beefmaster E6 - Certified by Beefmaster Breeders United to be at least 50% Beefmaster and can be as much as 100%. At least one of the parents must be registered as a purebred Beefmaster. • Golden Certified F1 – A female that is the progeny of two registered parents with one parent being registered Brahman resulting in a F1 cross (50% Brahman x 50% Bos Taurus). Must be issued a certificate of recordation form from ABBA which includes owner’s name, ownership date. -

Cattle Producer's Handbook
Western Beef Resource Committee Fourth Edition Cattle Producer’s Handbook Genetics Section 845 Breed Association Contact List J. Benton Glaze, Jr., University of Idaho Breed associations provide beef cattle producers a retain genetically superior animals for use in future variety of benefits and services. Breed associations work generations. To accomplish this task, producers must in the areas of breed promotion, marketing, member take advantage of available tools and resources, such education, performance recording, and performance as expected progeny differences (EPD). EPDs are an evaluation. While all services are important, one that evaluation of an animal’s genetic worth (value as a receives much attention is performance recording and parent). EPDs are reported in sire summaries, which are the evaluation of animals. published by several breed associations. To remain competitive in the beef cattle industry, Following is a list of breed associations and their producers must continually strive to identify and contact information. AMERIFAX BEEFALO Amerifax Cattle Association American Beefalo Association 400 N. Minnesota Ave. P.O. Box 295 P.O. Box 149 Benton City, WA 99320 Hastings, NE 68901 9824 E. YZ Ave. (402) 463-5289 Vicksburg, MI 49097 (800) 233-3256 ANGUS web: americanbeefalo.org American Angus Association 3201 Frederick Ave. BEEFMASTER St. Joseph, MO 64506 Beefmaster Breeders United (816) 383-5100 6800 Park Ten Blvd., Ste. 290W (816) 233-9703 fax San Antonio, TX 78213 web: www.angus.org (210) 732-3132 (210) 732-7711 fax BARZONA web: www.beefmasters.org Barzona Breeders Association of America 604 Cedar St. BLONDE D’AQUITAINE Adair, IA 50002 American Blonde D’Aquitaine Association (641) 745-9170 57 Friar Tuckway (641) 343-0927 fax Fyffe, AL 35971 web: www.barzona.com (256) 996-3142 web: www.blondecattle.org 845-1 BRAFORD GELBVIEH United Braford Breeders American Gelbvieh Association 638A N.