Taxonomic Status of Blackthroat Calliope Obscura and Firethroat C
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Flyer200206 Parent
THE, FLYE, R Volume 26,25, NumberNumber 6 6 tuneJune 20022002 NEXT MEETING After scientists declared the Gunnison Sage Grouse a new species two years ago, a wide spot on Gun- Summer is here and we won't have meeting until a nison County (Colorado) Road 887, has become an Wednesday, 18. It begin September will atl:30 international bird-watching sensation. Birders from p.m. in Room 117 Millington Hall, on the William around the ,world wait silently in the cold dark of a campus. The editors'also get a summer &Mary Colorado spring pre-dawn to hear the "Thwoomp! vacation so there will be no July Flyer, but "God Thwoomp! Thwoomp!" of a male Gunnison Sage the creek rise," there be an willing and if don't will Grouse preparing to mate. The noise comes from August issue. specialized air sacs on the bird's chest. And this is now one stop on a well-traveled 1,000 mile circuit being traveled by birders wanting to add this Gun- RAIN CURTAILS FIELD TRIP TO nison bird, plus the Chukar, the Greater Sage YORK RIVER STATE PARK Grouse, the White-tailed Ptarmigan, the Greater Chicken Skies were threatening and the wind was fierce at Prairie and the Lesser Prairie Chicken to the beginning of the trip to the York River State their lii'e lists. Park on May 18. Despite all of that, leader Tom It was not always like this. Prior to the two-year- Armour found some very nice birds before the rains ago decision by the Ornithological Union that this came flooding down. -

Rare Birds of California Now Available! Price $54.00 for WFO Members, $59.99 for Nonmembers
Volume 40, Number 3, 2009 The 33rd Report of the California Bird Records Committee: 2007 Records Daniel S. Singer and Scott B. Terrill .........................158 Distribution, Abundance, and Survival of Nesting American Dippers Near Juneau, Alaska Mary F. Willson, Grey W. Pendleton, and Katherine M. Hocker ........................................................191 Changes in the Winter Distribution of the Rough-legged Hawk in North America Edward R. Pandolfino and Kimberly Suedkamp Wells .....................................................210 Nesting Success of California Least Terns at the Guerrero Negro Saltworks, Baja California Sur, Mexico, 2005 Antonio Gutiérrez-Aguilar, Roberto Carmona, and Andrea Cuellar ..................................... 225 NOTES Sandwich Terns on Isla Rasa, Gulf of California, Mexico Enriqueta Velarde and Marisol Tordesillas ...............................230 Curve-billed Thrasher Reproductive Success after a Wet Winter in the Sonoran Desert of Arizona Carroll D. Littlefield ............234 First North American Records of the Rufous-tailed Robin (Luscinia sibilans) Lucas H. DeCicco, Steven C. Heinl, and David W. Sonneborn ........................................................237 Book Reviews Rich Hoyer and Alan Contreras ...........................242 Featured Photo: Juvenal Plumage of the Aztec Thrush Kurt A. Radamaker .................................................................247 Front cover photo by © Bob Lewis of Berkeley, California: Dusky Warbler (Phylloscopus fuscatus), Richmond, Contra Costa County, California, 9 October 2008, discovered by Emilie Strauss. Known in North America including Alaska from over 30 records, the Dusky is the Old World Warbler most frequent in western North America south of Alaska, with 13 records from California and 2 from Baja California. Back cover “Featured Photos” by © Kurt A. Radamaker of Fountain Hills, Arizona: Aztec Thrush (Ridgwayia pinicola), re- cently fledged juvenile, Mesa del Campanero, about 20 km west of Yecora, Sonora, Mexico, 1 September 2007. -

Does a Rival's Song Elicit Territorial Defense in a Tropical Songbird, The
ABC 2017, 4(2):146-153 Animal Behavior and Cognition https://doi.org/10.12966/abc.02.05.2017 ©Attribution 3.0 Unported (CC BY 3.0) Does a Rival’s Song Elicit Territorial Defense in a Tropical Songbird, the Pied Bush Chat (Saxicola caprata)? Navjeevan Dadwal1* and Dinesh Bhatt1 1Gurukula Kangri University, Haridwar, Uttarakhand, India *Corresponding author (Email:[email protected]) Citation – Dadwal, N., & Bhatt, D. (2017). Does a rival’s song elicit territorial defense in a tropical songbird, the Pied Bush Chat (Saxicola caprata)? Animal Behavior and Cognition, 4(2), 146–153. https://doi.org/10.12966/ abc.02.05.2017 Abstract -The purpose of bird song and the way in which it is delivered has been argued to be adapted mainly for territorial defense. We performed a field experiment with the combination of playbacks and a model to test how much song actually relates to increased territorial defense in the territorial tropical songbird, the Pied Bush Chat, during breeding season (Feb–May, 2015) at Haridwar, Himalayan Foothills, India. As expected, the results of the experiment indicated that song was the major cue used by territory holders to cope with rival intrusions. The song rate was particularly escalated during simulated territorial interactions when the model was presented with a playback song of conspecifics. Behaviors such as restlessness (perch change), the height of perch, and distance from the model appeared to be of relatively lesser importance. To our knowledge, no avian species from the Indian subcontinent has been studied to provide evidence that song can escalate aggressive response by a territory owner. -
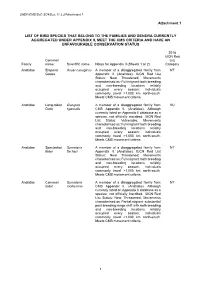
Attachment 1 LIST of BIRD SPECIES THAT BELONG to the FAMILIES
UNEP/CMS/ScC-SC4/Doc.11.3.2/Attachment 1 Attachment 1 LIST OF BIRD SPECIES THAT BELONG TO THE FAMILIES AND GENERA CURRENTLY AGGREGATED UNDER APPENDIX II, MEET THE CMS CRITERIA AND HAVE AN UNFAVOURABLE CONSERVATION STATUS 2018 IUCN Red Common List Family name Scientific name Notes for Appendix II (Sheets 1 or 2) Category Anatidae Emperor Anser canagicus A member of a disaggregated family from NT Goose Appendix II. (Anatidae). IUCN Red List Status: Near Threatened; Movements characterised as: Full migrant: both breeding and non-breeding locations reliably occupied every season; individuals commonly travel >1,000 km north-south. Meets CMS movement criteria. Anatidae Long-tailed Clangula A member of a disaggregated family from VU Duck hyemalis CMS Appendix II. (Anatidae). Although currently listed on Appendix II database as a species, not officially inscribed. IUCN Red List Status: Vulnerable; Movements characterised as: Full migrant: both breeding and non-breeding locations reliably occupied every season; individuals commonly travel >1,000 km north-south. Meets CMS movement criteria. Anatidae Spectacled Somateria A member of a disaggregated family from NT Eider fischeri Appendix II. (Anatidae). IUCN Red List Status: Near Threatened; Movements characterised as: Full migrant: both breeding and non-breeding locations reliably occupied every season; individuals commonly travel >1,000 km north-south. Meets CMS movement criteria. Anatidae Common Somateria A member of a disaggregated family from NT Eider mollissima CMS Appendix II. (Anatidae). Although currently listed on Appendix II database as a species, not officially inscribed. IUCN Red List Status: Near Threatened; Movements characterised as: Partial migrant: substantial post-breeding range shift with both breeding and non-breeding locations reliably occupied every season; individuals commonly travel >1,000 km north-south. -

Birdlife International for the Input of Analyses, Technical Information, Advice, Ideas, Research Papers, Peer Review and Comment
UNEP/CMS/ScC16/Doc.10 Annex 2b CMS Scientific Council: Flyway Working Group Reviews Review 2: Review of Current Knowledge of Bird Flyways, Principal Knowledge Gaps and Conservation Priorities Compiled by: JEFF KIRBY Just Ecology Brookend House, Old Brookend, Berkeley, Gloucestershire, GL13 9SQ, U.K. June 2010 Acknowledgements I am grateful to colleagues at BirdLife International for the input of analyses, technical information, advice, ideas, research papers, peer review and comment. Thus, I extend my gratitude to my lead contact at the BirdLife Secretariat, Ali Stattersfield, and to Tris Allinson, Jonathan Barnard, Stuart Butchart, John Croxall, Mike Evans, Lincoln Fishpool, Richard Grimmett, Vicky Jones and Ian May. In addition, John Sherwell worked enthusiastically and efficiently to provide many key publications, at short notice, and I’m grateful to him for that. I also thank the authors of, and contributors to, Kirby et al. (2008) which was a major review of the status of migratory bird species and which laid the foundations for this work. Borja Heredia, from CMS, and Taej Mundkur, from Wetlands International, also provided much helpful advice and assistance, and were instrumental in steering the work. I wish to thank Tim Jones as well (the compiler of a parallel review of CMS instruments) for his advice, comment and technical inputs; and also Simon Delany of Wetlands International. Various members of the CMS Flyway Working Group, and other representatives from CMS, BirdLife and Wetlands International networks, responded to requests for advice and comment and for this I wish to thank: Olivier Biber, Joost Brouwer, Nicola Crockford, Carlo C. Custodio, Tim Dodman, Roger Jaensch, Jelena Kralj, Angus Middleton, Narelle Montgomery, Cristina Morales, Paul Kariuki Ndang'ang'a, Paul O’Neill, Herb Raffaele and David Stroud. -

Disaggregation of Bird Families Listed on Cms Appendix Ii
Convention on the Conservation of Migratory Species of Wild Animals 2nd Meeting of the Sessional Committee of the CMS Scientific Council (ScC-SC2) Bonn, Germany, 10 – 14 July 2017 UNEP/CMS/ScC-SC2/Inf.3 DISAGGREGATION OF BIRD FAMILIES LISTED ON CMS APPENDIX II (Prepared by the Appointed Councillors for Birds) Summary: The first meeting of the Sessional Committee of the Scientific Council identified the adoption of a new standard reference for avian taxonomy as an opportunity to disaggregate the higher-level taxa listed on Appendix II and to identify those that are considered to be migratory species and that have an unfavourable conservation status. The current paper presents an initial analysis of the higher-level disaggregation using the Handbook of the Birds of the World/BirdLife International Illustrated Checklist of the Birds of the World Volumes 1 and 2 taxonomy, and identifies the challenges in completing the analysis to identify all of the migratory species and the corresponding Range States. The document has been prepared by the COP Appointed Scientific Councilors for Birds. This is a supplementary paper to COP document UNEP/CMS/COP12/Doc.25.3 on Taxonomy and Nomenclature UNEP/CMS/ScC-Sc2/Inf.3 DISAGGREGATION OF BIRD FAMILIES LISTED ON CMS APPENDIX II 1. Through Resolution 11.19, the Conference of Parties adopted as the standard reference for bird taxonomy and nomenclature for Non-Passerine species the Handbook of the Birds of the World/BirdLife International Illustrated Checklist of the Birds of the World, Volume 1: Non-Passerines, by Josep del Hoyo and Nigel J. Collar (2014); 2. -

Thailand Highlights 14Th to 26Th November 2019 (13 Days)
Thailand Highlights 14th to 26th November 2019 (13 days) Trip Report Siamese Fireback by Forrest Rowland Trip report compiled by Tour Leader: Forrest Rowland Trip Report – RBL Thailand - Highlights 2019 2 Tour Summary Thailand has been known as a top tourist destination for quite some time. Foreigners and Ex-pats flock there for the beautiful scenery, great infrastructure, and delicious cuisine among other cultural aspects. For birders, it has recently caught up to big names like Borneo and Malaysia, in terms of respect for the avian delights it holds for visitors. Our twelve-day Highlights Tour to Thailand set out to sample a bit of the best of every major habitat type in the country, with a slight focus on the lush montane forests that hold most of the country’s specialty bird species. The tour began in Bangkok, a bustling metropolis of winding narrow roads, flyovers, towering apartment buildings, and seemingly endless people. Despite the density and throng of humanity, many of the participants on the tour were able to enjoy a Crested Goshawk flight by Forrest Rowland lovely day’s visit to the Grand Palace and historic center of Bangkok, including a fun boat ride passing by several temples. A few early arrivals also had time to bird some of the urban park settings, even picking up a species or two we did not see on the Main Tour. For most, the tour began in earnest on November 15th, with our day tour of the salt pans, mudflats, wetlands, and mangroves of the famed Pak Thale Shore bird Project, and Laem Phak Bia mangroves. -

South Africa Mega Birding Tour I 6Th to 30Th January 2018 (25 Days) Trip Report
South Africa Mega Birding Tour I 6th to 30th January 2018 (25 days) Trip Report Aardvark by Mike Bacon Trip report compiled by Tour Leader: Wayne Jones Rockjumper Birding Tours View more tours to South Africa Trip Report – RBT South Africa - Mega I 2018 2 Tour Summary The beauty of South Africa lies in its richness of habitats, from the coastal forests in the east, through subalpine mountain ranges and the arid Karoo to fynbos in the south. We explored all of these and more during our 25-day adventure across the country. Highlights were many and included Orange River Francolin, thousands of Cape Gannets, multiple Secretarybirds, stunning Knysna Turaco, Ground Woodpecker, Botha’s Lark, Bush Blackcap, Cape Parrot, Aardvark, Aardwolf, Caracal, Oribi and Giant Bullfrog, along with spectacular scenery, great food and excellent accommodation throughout. ___________________________________________________________________________________ Despite havoc-wreaking weather that delayed flights on the other side of the world, everyone managed to arrive (just!) in South Africa for the start of our keenly-awaited tour. We began our 25-day cross-country exploration with a drive along Zaagkuildrift Road. This unassuming stretch of dirt road is well-known in local birding circles and can offer up a wide range of species thanks to its variety of habitats – which include open grassland, acacia woodland, wetlands and a seasonal floodplain. After locating a handsome male Northern Black Korhaan and African Wattled Lapwings, a Northern Black Korhaan by Glen Valentine -

Namibia & the Okavango
Pel’s Fishing Owl - a pair was found on a wooded island south of Shakawe (Jan-Ake Alvarsson) NAMIBIA & THE OKAVANGO 21 SEPTEMBER – 8 OCTOBER 2017 LEADER: STEVE BRAINE For most of the country the previous three years drought had been broken and although too early for the mi- grants we did however do very well with birding generally. We searched and found all the near endemics as well as the endemic Dune Lark. Besides these we also had a new write-in for the trip! In the floodplains after observing a wonderful Pel’s Fishing Owl we travelled down a side channel of the Okavango River to look for Pygmy Geese, we were lucky and came across several pairs before reaching a dried-out floodplain. Four birds flew out of the reedbeds and looked rather different to the normal weavers of which there were many, a closer look at the two remaining birds revealed a beautiful pair of Cuckoo Finches. These we all enjoyed for a brief period before they followed the other birds which had now disappeared into the reedbeds. Very strong winds on three of the birding days made birding a huge challenge to say the least after not finding the rare and difficult Herero Chat we had to make alternate arrangements at another locality later in the trip. The entire tour from the Hosea Kutako International Airport outside the capital Windhoek and returning there nineteen days later delivered 375 species. Out of these, four birds were seen only by the leader, a further three species were heard but not seen. -

SICHUAN (Including Northern Yunnan)
Temminck’s Tragopan (all photos by Dave Farrow unless indicated otherwise) SICHUAN (Including Northern Yunnan) 16/19 MAY – 7 JUNE 2018 LEADER: DAVE FARROW The Birdquest tour to Sichuan this year was a great success, with a slightly altered itinerary to usual due to the closure of Jiuzhaigou, and we enjoyed a very smooth and enjoyable trip around the spectacular and endemic-rich mountain and plateau landscapes of this striking province. Gamebirds featured strongly with 14 species seen, the highlights of them including a male Temminck’s Tragopan grazing in the gloom, Chinese Monal trotting across high pastures, White Eared and Blue Eared Pheasants, Lady Amherst’s and Golden Pheasants, Chinese Grouse and Tibetan Partridge. Next were the Parrotbills, with Three-toed, Great and Golden, Grey-hooded and Fulvous charming us, Laughingthrushes included Red-winged, Buffy, Barred, Snowy-cheeked and Plain, we saw more Leaf Warblers than we knew what to do with, and marvelled at the gorgeous colours of Sharpe’s, Pink-rumped, Vinaceous, Three-banded and Red-fronted Rosefinches, the exciting Przevalski’s Finch, the red pulse of Firethroats plus the unreal blue of Grandala. Our bird of the trip? Well, there was that Red Panda that we watched for ages! 1 BirdQuest Tour Report: Sichuan Including Northern Yunnan 2018 www.birdquest-tours.com Our tour began with a short extension in Yunnan, based in Lijiang city, with the purpose of finding some of the local specialities including the rare White-speckled Laughingthrush, which survives here in small numbers. Once our small group had arrived in the bustling city of Lijiang we began our birding in an area of hills that had clearly been totally cleared of forest in the fairly recent past, with a few trees standing above the hillsides of scrub. -

Siberian Blue Robin Larvivora Cyane from the Barak Valley of Assam with a Status Update for India
See discussions, stats, and author profiles for this publication at: https://www.researchgate.net/publication/338137338 Siberian Blue Robin Larvivora cyane from the Barak Valley of Assam with a status update for India Article in Indian BIRDS · December 2019 CITATIONS READS 0 7 2 authors: Rejoice Gassah Vijay Anand Ismavel Makunda Christian Hospital 4 PUBLICATIONS 0 CITATIONS 22 PUBLICATIONS 12 CITATIONS SEE PROFILE SEE PROFILE Some of the authors of this publication are also working on these related projects: Neglected health problems in rural India View project Biodiversity Documentation View project All content following this page was uploaded by Vijay Anand Ismavel on 24 December 2019. The user has requested enhancement of the downloaded file. Correspondence 123 peninsular India (Grimmett et al. 2011; Rasmussen & Anderton minY=11.103753465762485&env.maxX=93.01342361450202&env.maxY=12.31963 2012; eBird 2019). This species is a common winter visitor to 3103994705&zh=true&gp=true&ev=Z&mr=on&bmo=1&emo=1&yr=cur&byr=2019 Myanmar, Thailand, Laos, and Tonkin (Robson 2008). &eyr=2019. [Accessed on: 19 January 2019.] DeCandido, R., Subedi, T., Siponen, M., Sutasha, K., Pierce, A., Nualsri, C., & Round, P. D., 2013. Flight identification of Milvus migrans lineatus ‘Black-eared’ Kite and Milvus migrans govinda ‘Pariah’ Kite in Nepal and Thailand. BirdingASIA 20: 32–36. Grimmett, R., Inskipp, C., & Inskipp, T., 2011. Birds of the Indian Subcontinent. 2nd ed. London: Oxford University Press & Christopher Helm. Pp. 1–528. Rasmussen, P. C., & Anderton, J. C., 2012. Birds of South Asia: the Ripley guide: field guide. 2nd ed. Washington, D.C. -

First Images in the Wild of Blackthroat Luscinia Obscura, Asia's Most
BirdingASIA 15 (2011): 17–19 17 LITTLE-KNOWN ASIAN BIRD First images in the wild of Blackthroat Luscinia obscura, Asia’s most enigmatic robin WEI QIAN & HE YI Background probably a female from Baihe Nature Reserve, The Blackthroat Luscinia obscura is known to breed Sichuan, on 3 June 2007 (Anderson 2007). only in the mountains of south-west China, where Additionally, a few captive birds, assumed to have there are a few scattered records from Sichuan, been caught in the vicinity of Chengdu, have Gansu and Shaanxi, together with even fewer appeared in Chengdu bird market (Wang 2004). presumed non-breeding records from Yunnan All other records are from Thailand and presumed (southern China) and northern Thailand. It is an to be either wintering birds or passage migrants extremely poorly known species classified as (BirdLife International 2001). Vulnerable because it is inferred to have a small, declining population as a result of destruction of Observations temperate forest within its breeding area as well On the morning of 2 May 2011, we visited Sichuan as habitat loss in its likely wintering areas (BirdLife University to observe and photograph migrating International 2001, 2011). birds in a small patch of wood and shrubbery in Since its description in 1891, there have only the south-eastern corner of the campus. At about been nine records from China, including some 09h20 a bird abruptly flew into the shrubbery, uncertain ones. These records comprise one each which we quickly located and identified as a male from Ganshu and Shaanxi, two from Yunnan, Blackthroat Luscinia obscura. We continued to presumed to be during migration, and four from observe it until about 17h20 when we left and Sichuan, all detailed in BirdLife International obtained what we believe to be the first images of (2001), and a recent sight record of a male and this species in the wild (Plates 1 & 2).