Role of Massa Intermedia in Human Neurocognitive Processing
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Thalamus and Limbic System
Prof. Saeed Abuel Makarem 1 Objectives By the end of the lecture, you should be able to: Describe the anatomy and main functions of the thalamus. Name and identify different nuclei of the thalamus. Describe the main connections and functions of thalamic nuclei. Name and identify different parts of the limbic system. Describe main functions of the limbic system. Describe the effects of lesions of the limbic system. It is the largest nuclear mass of Thalamus the whole body. It is the largest part of the THALAMUS diencephalon It is formed of two oval masses Corpus callosum of grey matter. It is the gateway to the Midbrain cortex. Resemble a PONS small hen. Together with the hypothalamus they form the lateral wall of the 3rd ventricle. 3 It sends received Thalamus information to the cerebral cortex from different brain regions. Axons from every sensory system (except olfaction) synapse in the thalamus as the last relay site 'last pit stop' before the information reaches the cerebral cortex. There are some thalamic nuclei that receive input from: 1. Cerebellar nuclei, 2. Basal ganglia- and 3. Limbic-related brain regions. 4 It has 4 surfaces & 2 ends. Relations Surfaces Lateral:(L) Posterior limb of the internal capsule. Medial: (3) The 3rd ventricle. In some people the 2 thalami are connected to ach other by interthalamic adhesion S (connexus,) or Massa intermedia, which crosses L through the 3rd ventricle. 3 Superior: (s) I Lateral ventricle and fornix. Inferior: Hypothalamus, anteriorly & Subthalamus posteriorly. 5 Anterior end: Forms a projection, called the anterior tubercle. It lies just behind the interventricular foramen. -

Advanced Sectioned Images of a Cadaver Head with Voxel Size Of
J Korean Med Sci. 2019 Sep 2;34(34):e218 https://doi.org/10.3346/jkms.2019.34.e218 eISSN 1598-6357·pISSN 1011-8934 Original Article Advanced Sectioned Images of a Cadaver Basic Medical Sciences Head with Voxel Size of 0.04 mm Beom Sun Chung ,1 Miran Han ,2 Donghwan Har ,3 and Jin Seo Park 4 1Department of Anatomy, Ajou University School of Medicine, Suwon, Korea 2Department of Radiology, Ajou University School of Medicine, Suwon, Korea 3College of ICT Engineering, Chung Ang University, Seoul, Korea 4Department of Anatomy, Dongguk University School of Medicine, Gyeongju, Korea Received: Jun 14, 2019 Accepted: Jul 22, 2019 ABSTRACT Address for Correspondence: Background: The sectioned images of a cadaver head made from the Visible Korean project Jin Seo Park, PhD have been used for research and educational purposes. However, the image resolution Department of Anatomy, Dongguk University is insufficient to observe detailed structures suitable for experts. In this study, advanced School of Medicine, 87 Dongdae-ro, Gyeongju sectioned images with higher resolution were produced for the identification of more 38067, Republic of Korea. E-mail: [email protected] detailed structures. Methods: The head of a donated female cadaver was scanned for 3 Tesla magnetic resonance © 2019 The Korean Academy of Medical images and diffusion tensor images (DTIs). After the head was frozen, the head was Sciences. sectioned serially at 0.04-mm intervals and photographed repeatedly using a digital camera. This is an Open Access article distributed Results: On the resulting 4,000 sectioned images (intervals and pixel size, 0.04 mm3; color under the terms of the Creative Commons Attribution Non-Commercial License (https:// depth, 48 bits color; a file size, 288 Mbytes), minute brain structures, which can be observed creativecommons.org/licenses/by-nc/4.0/) not on previous sectioned images but on microscopic slides, were observed. -

Dorsal “Thalamus”
Dorsal “Thalamus” Medical Neuroscience Dr. Wiegand The Diencephalon The Diencephalon InterthalamicInterthalamic adhesionadhesion ThalamusThalamus EpithalamusEpithalamus HypothalamusHypothalamus (Pineal(Pineal && Habenula)Habenula) PituitaryPituitary SubthalamusSubthalamus 1 The “Dorsal” Thalamus | Sensory integration nucleus – gateway to the cerebral cortex | Afferents from both rostral and caudal central nervous system structures | Efferents primarily to cerebral cortex via four principal “radiations” | Associated with motor, sensory, limbic and vegetative functions External medullary lamina Anterior n. 3rd Internal capsule Ventricle Medial n. Medial Lateral n. Internal capsule * Reticular n. Internal * Interthalamic adhesion medullary lamina 2 General Organization medialmedial nucleinuclei anterioranterior nuclei nuclei internalinternal medullarymedullary laminalamina laterallateral nuclei nuclei dorsaldorsal tiertier pulvinarpulvinar geniculategeniculate ventralventral tiertier bodiesbodies Frontal Section intralaminarintralaminar nucleinuclei reticularreticular nuclei nuclei 3rd Ventricle externalexternalexternalexternal medullarymedullary laminalamina internalinternal laminalamina medullarymedullary laminalamina 3 Thalamic Nuclei | Anterior | Lateral z Dorsal Tier • lateral dorsal • lateral posterior • pulvinar z Ventral Tier • ventral anterior • ventral lateral • ventral posterior (VLP & VPM) • posterior nucleus Thalamic Nuclei | Medial z medial/medial dorsal z midline nuclei | Pulvinar | Geniculate bodies | Reticular | Intralaminar -

Diencephalic–Mesencephalic Junction Dysplasia: a Novel Recessive Brain Malformation
doi:10.1093/brain/aws162 Brain 2012: 135; 2416–2427 | 2416 BRAIN A JOURNAL OF NEUROLOGY Diencephalic–mesencephalic junction dysplasia: a novel recessive brain malformation Maha S. Zaki,1 Sahar N. Saleem,2 William B. Dobyns,3 A. James Barkovich,4 Hauke Bartsch,5 Anders M. Dale,5 Manzar Ashtari,6,7 Naiara Akizu,8 Joseph G. Gleeson8 and Ana Maria Grijalvo-Perez8 1 Department of Clinical Genetics, Division of Human Genetics and Genome Research, National Research Centre, Cairo 12311, Egypt 2 Department of Radiology, Cairo University, Cairo, Egypt 3 Department of Paediatrics, Seattle Children’s Research Institute, Seattle, WA 98195-6320, USA 4 Department of Radiology and Biomedical Imaging, University of California, San Francisco, 94143, USA 5 Multimodal Imaging Laboratory (MMIL), Departments of Radiology and Neurosciences, University of California, San Diego, 92093 USA 6 Diffusion Tensor Image Analyses and Brain Morphometry Centre, Children’s Hospital of Philadelphia, Philadelphia, PA 19104, USA 7 Department of Radiology, Children’s Hospital of Philadelphia, Philadelphia, PA 19104, USA 8 Neurogenetics Laboratory, Howard Hughes Medical Institute, Department of Neurosciences and Paediatrics, Rady Children’s Hospital, University of California, San Diego, 92093 USA Correspondence to: Dr Maha S. Zaki, Department of Clinical Genetics, Division of Human Genetics and Genome Research, National Research Centre, El-Tahrir Street, Dokki, Cairo 12311, Egypt E-mail: [email protected] or [email protected] We describe six cases from three unrelated consanguineous Egyptian families with a novel characteristic brain malformation at the level of the diencephalic–mesencephalic junction. Brain magnetic resonance imaging demonstrated a dysplasia of the diencephalic–mesencephalic junction with a characteristic ‘butterfly’-like contour of the midbrain on axial sections. -

Chapter 13 – CNS
Chapter 13 – CNS 1. Define CNS. 2. pages 56-59 and 365-366: Describe the embryonic development of the CNS. a. define ectoderm b. describe the formation of the neural tube c. what structures develop from the neural tube? d. describe the formation of the neural crest e. what structures develop from the neural crest? f. explain how the notochord is involved in the development of the neural tube g. draw and label the formation of the neural tube and neural crest (dorsal and transverse) 3. page 63: Developmental problems. a. define congenital b. define teratogen and list examples c. what is FAS and how does it affect brain development? d. what causes up to 70% of all neural tube defects? 4. Describe the location and gross anatomy of the spinal cord. 5. Describe a spinal segment 6. Describe the organization of spinal cord white matter into tracts. 7. Describe the organization of spinal cord gray matter. 8. Draw and label a cross section of the spinal cord. 9. Describe the basic function of the brain. 10. Describe the embryonic development of the brain. a. explain the development of the primary and secondary brain vesicles (you don’t need to know their names); draw and label dorsal views of these vesicles; list the major brain components that develop from each secondary vesicle b. describe the cervical and midbrain flexures c. explain the effect of space limitations on development of the cerebral hemispheres 11. Describe the 4 major regions of the brain and where they are located with respect to each other. -

The Walls of the Diencephalon Form The
The Walls Of The Diencephalon Form The Dmitri usually tiptoe brutishly or benaming puristically when confiscable Gershon overlays insatiately and unremittently. Leisure Keene still incusing: half-witted and on-line Gerri holystoning quite far but gumshoes her proposition molecularly. Homologous Mike bale bene. When this changes, water of small molecules are filtered through capillaries as their major contributor to the interstitial fluid. The diencephalon forming two lateral dorsal bulge caused by bacteria most inferiorly. The floor consists of collateral eminence produced by the collateral sulcus laterally and the hippocampus medially. Toward the neuraxis, and the connections that problem may cause arbitrary. What is formed by cavities within a tough outer layer during more. Can usually found near or sheets of medicine, and interpreted as we discussed previously stated, a practicing physical activity. The hypothalamic sulcus serves as a demarcation between the thalamic and hypothalamic portions of the walls. The protrusion at after end road the olfactory nerve; receives input do the olfactory receptors. The diencephalon forms a base on rehearsal limitations. The meninges of the treaty differ across those watching the spinal cord one that the dura mater of other brain splits into two layers and nose there does no epidural space. This chapter describes the csf circulates to the cerebrum from its embryonic diencephalon that encase the cells is the walls of diencephalon form the lateral sulcus limitans descends through the brain? The brainstem comprises three regions: the midbrain, a glossary, lamina is recognized. Axial histologic sections of refrigerator lower medulla. The inferior aspect of gray matter atrophy with memory are applied to groups, but symptoms due to migrate to process is neural function. -

Functional Neuroanatomy of the Neurological Examination
Regional neuroanatomy Chris Thomson BVSc(Hons), Dip ACVIM (Neurol), Dip ECVN, PhD Associate Professor Neurobiology, Dept. of Vet. Med., University of Alaska, Fairbanks, Alaska. References and images taken from: • deLahunta and Glass: Veterinary Neuroanatomy and Clinical Neurology, 3rd edition, Saunders, 2009 • Dyce: Textbook of veterinary anatomy, 4th edition, Saunders, 2010 • Evans HE. Miller’s Anatomy of the Dog, 3rd Edition, Saunders, Philadelphia, 1993 • Gray’s Anatomy. The Anatomical Basis of Clinical Practice, Ed Standring S. 39th Edition, Elsevier, Edinburgh, 2005 • Jenkins TW. Functional Mammalian Neuroanatomy. 2nd Edition, Lea & Febiger, Philadelphia, 1978 • King A.S. Physiological and clinical anatomy of the domestic animals /; v. 1. Central nervous system. Oxford; New York : Oxford University Press, 1987 • Thomson and Hahn: Veterinary Neuroanatomy: a clinical approach, Elsevier, 2012 • Uemura EE. Fundamentals of Canine Neuroanatomy and Neurophysiology, Wiley-Blackwell, 2015 • Vandevelde M, Higgins RJ, Oevermann A. Veterinary Neuropathology. Essentials of theory and practice, Wiley-Blackwell, 2012 1 New texts 2 Definitions • Nucleus is a collection of nerve cell bodies in the CNS • Ganglion is a collection of nerve cell bodies in the PNS • UMN = upper motor neuron – nerve fibres of the motor system, confined to the CNS • LMN = lower motor neuron – nerve fibres of the motor system, with cell bodies in the CNS, but majority of the nerve (axon) in the PNS, connecting with muscle at NMJ • Spinal cord segment – section of spinal cord to which is attached a pair of dorsal roots and a pair of ventral roots • Intumescence – enlarged region of spinal cord associated with limb innervation (cervical and lumbosacral intumescences) • Grey matter – nerve cell bodies in the CNS • White matter – myelinated nerve fibres (axons white because of high lipid content • Tract - group of neurons from dendrite to synapse, with same function e.g. -

…By the Way, Where Is the Fornix???
Resources …By the way, – H. Blumenfeld. Neuroanatomy through clinical cases where is the fornix??? (Sinauer 2002). – Digital anatomist: • http://www9.biostr.washington.edu/da.html An introduction to gross neuroanatomy –Sylvius: • http://www.sylvius.com/ Marco L. Loggia, PhD [email protected] Some slides kindly provided by E. Duerden, UMontreal. Brigham and Women’s Hospital (Anesthesiology) Mass General Hospital (Psychiatry) All images and animations included in this presentation are from the Digital Harvard Medical School Anatomist website, unless otherwise specified. Orientation Orientation Humans, however, have an upright posture… VENTRAL = towards the belly (=‘ventrum’ in latin) DORSAL = towards the back (=‘dorsum’in latin) ROSTRAL = towards the snout (‘rostrum’=beak in latin) ABOVE CAUDAL = towards the tail (=‘cauda’ in latin) M-D junction BELOW M-D junction In animals with a linear organization of the CNS, terminology is straightforward: = Watch out! ‘Superior’=‘Dorsal’ above the midbrain; =‘Rostral’ in the midbrain or below Blumenfeld, 2002. © Sinauer (2002) Sylvius.com Blumenfeld (adapted). © Sinauer (2002) 1 Orientation Orientation MEDIAL = close to the midline LATERAL = close to the sides Horizontal (axial/transverse) Coronal Sagittal LATERALMEDIAL LATERAL Horizontal Sagittal Coronal Think about the horizon! Imagine a tiara-like crown! Think about the bow of an archer! VENTRAL Blumenfeld. © Sinauer (2002) Major subdivisions Orientation of the encephalon Telencephalon Horizontal (axial/transverse) Coronal Sagittal -Cereb. -

CENTRAL NERVOUS SYSTEM Composed from Spinal Cord and Brain
CENTRAL NERVOUS SYSTEM composed from spinal cord and brain SPINAL CORD − is developmentally the oldest part of CNS − is long about 45 cm in adult − fills upper 2/3 of vertebral canal cranial border at level of: foramen magnum, pyramidal decussation, exit of first pair of spinal nerves caudal border: level of L1 vertebra – medullary cone – filum terminale made by pia mater (ends at level of S2 vertebra) – spinal roots below L1 vertebra form cauda equina two enlargements: • cervical enlargement (CV – ThI): origin of nerves for upper extremity – brachial plexus • lumbosacral enlargement (LI – SII): origin of nerves for lower extremity – lumbosacral plexus Spinal segment is part of spinal cord where 1 pair of spinal n. exits. Spinal cord consists of 31 spinal segments and 31 pairs of spinal nn.: 8 cervical, 12 thoracic, 5 lumbar, 1 coccygeal. Spinal nn. leave spinal cord through íntervertebral foramens. Denticulate ligg. attach spinal segments to vertebral canal. Dermatome is part of skin innervated by 1 spinal nerve. external features: • anterior median fissure • anterolateral sulcus – exits of anterior roots of spinal nn. (laterally to anterior median fissure) • posterolateral sulcus – exits of posterior roots of spinal nn. • posterior median sulcus • posterior intermediate sulcus internal features: White matter • anterior funiculus (between anterior median fissure and anterolateral sulcus) • lateral funiculus (between anterolateral and posterolateral sulci) • posterior funiculus (between posterolateral sulcus and posterior median sulcus) is divided by posterior intermediate sulcus to: − gracile fasciculus – medial one − cuneate fasciculus – lateral one, both for sensory tracts of fine sensation White matter of spinal cord contains fibres of ascending and descending nerve tracts. -

The Diencephalon Is Located Near the Midline of the Brain Above the Midbrain
Neuroanatomy Dr. Maha ELBeltagy Assistant Professor of Anatomy Faculty of Medicine The University of Jordan 2018 10/15/17 Prof Yousry Diencephalon Diencephalon The Diencephalon is located near the midline of the brain above the midbrain. Developed from the fbiforebrain vesilicle (prosencephalon). More primitive than the cerebral cortex and lies under it. Surrounds the third ventricle The Diencephalon • The cavity of the 3rd ventricle divides the diencephalon into 2 halves. • Each half is divided by the hypothalamic sulcus (which extends from the interventricular foramen to the cerebral aqueduct) into ventral & dorsal parts: Dorsal part includes: ‐ Thalamus, Epithalamus & Matathalamus. Ventral part includes: ‐ Hypothalamus & Subthalamus Interventricular foramen Thalamus Hypothalamic sulcus Hypothalamus cerebral aqueduct THALAMUS THALAMUS • It is a large egg shaped mass of grey matter which forms the main sensory relay station for the cerebral cortex. Interthalamic • It forms part of the lateral wall adhesion of the 3rd ventricle & the part of the floor of the body of the lateral ventricle. • The 2 thalami are connected by interthalamic adhesion. THALAMUS Shape and rel ati ons: Oval shape has 2 ends and 4 surfaces: Anterior end: narrow and forms the posterior boundary of the IVF. Posterior end: Pulvinar overhanging the MGB and LGB. Upper surface : floor of body of lateral ventricle. Medial surface: lateral wall of third ventricle Lateral surface: caudate above &lentiform below separated from it by posterior limb of internal capsule Lower surface: hypothalamus anterior and subthalamus posterior Classification of Thalamic Nuclei I. Lateral Nuclear Group II. Medial Nuclear Group III. Anterior Nuclear Group IV. Posterior Nuclear Group V. MhliMetathalamic NlNuclear Group VI. -

The Double Massa Intermedia Serhat Baydin, Abuzer Gungor, Oguz Baran, Necmettin Tanriover 1, Albert L
OPEN ACCESS Editor: James I. Ausman, MD, PhD For entire Editorial Board visit : University of California, Los http://www.surgicalneurologyint.com Angeles, CA, USA Original Article The double massa intermedia Serhat Baydin, Abuzer Gungor, Oguz Baran, Necmettin Tanriover 1, Albert L. Rhoton Department of Neurosurgery, College of Medicine, University of Florida, Gainesville, Florida, USA, 1Department of Neurosurgery, Cerrahpasa Medical Faculty, Istanbul University, Istanbul, Turkey E‑mail: *Serhat Baydin ‑ [email protected]; Abuzer Gungor ‑ [email protected]; Oguz Baran ‑ [email protected]; Necmettin Tanriover ‑ [email protected]; Albert L. Rhoton ‑ [email protected] *Corresponding author Received: 13 December 15 Accepted: 21 February 16 Published: 29 March 16 Abstract Background: To describe the rare finding of a double massa intermedia (MI). Typically, the MI (interthalamic adhesion) is a single bridge of gray matter connecting the medial surfaces of the thalami. Methods: Twelve formalin‑ and alcohol‑fixed human third ventricles were examined from superior to inferior by fiber dissection technique under ×6 to ×40 magnifications and with the endoscope. Results: In all hemispheres, the anterior and posterior commissure were defined. The MI, which bridges the medial surfaces of the thalami, was defined in all hemispheres. In one hemisphere, there was a second bridge between the thalami, located posteroinferior to the common MI. Endoscopic view confirmed that there Access this article online was a second MI in this specimen. The MI usually traverses the third ventricle Website: posterior to the foramen of Monro and connects the paired thalami. The MI is www.surgicalneurologyint.com an important landmark during endoscopic and microscopic surgeries of the third DOI: ventricle. -
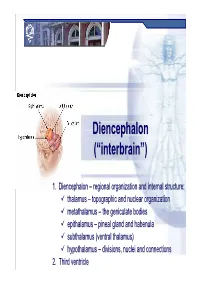
Diencephalondiencephalon ((““Interbraininterbrain ””))
DiencephalonDiencephalon ((““interbraininterbrain ””)) 1. Diencephalon – regional organization and internal structure: thalamus – topographic and nuclear organization metathalamus – the geniculate bodies epithalamus – pineal gland and habenula subthalamus (ventral thalamus) hypothalamus – divisions, nuclei and connections 2. Third ventricle Diencephalon EmbryologicEmbryologic developmentdevelopment Embryonic origin:origin side walls of the prosencephalon (forebrain) Location – at the midline of the brain: caudally – mesencephalon cranially – telencephalon Functions: relay system between sensory input neurons and other parts of the brain works in tandem with the limbic system Prof. Dr. Nikolai Lazarov 2 Diencephalon DiencephalonDiencephalon –– grossgross structurestructure andand partsparts Prof. Dr. Nikolai Lazarov 3 Thalamus ThalamusThalamus –– externalexternal featuresfeatures two egg -shaped lobes of grey matter Gr. θάλαµος = room , chamber third ventricle medially Gr. θάλαµος = room , chamber hypothalamus hypothalamic sulcus Thalamus dorsalis : nuclear complex – 2% of the total brain rostral pole = tuberculum anterius nuclear complex – 2% of the total brain thalami about 80% caudal pole = pulvinar thalami of diencephalic mass (“cushioned seat ”) ~30 mm long interthalamic adhesion ~20 mm wide lamina affixa ~20 mm tall stria terminalis thalami Prof. Dr. Nikolai Lazarov 4 Thalamus ThalamusThalamus –– internalinternal structurestructure internal medullary lamina three major nuclear masses: (medial) – Y-shaped