Robert Bowen Paleotemperature
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

This Article Appeared in a Journal Published by Elsevier. the Attached
This article appeared in a journal published by Elsevier. The attached copy is furnished to the author for internal non-commercial research and education use, including for instruction at the authors institution and sharing with colleagues. Other uses, including reproduction and distribution, or selling or licensing copies, or posting to personal, institutional or third party websites are prohibited. In most cases authors are permitted to post their version of the article (e.g. in Word or Tex form) to their personal website or institutional repository. Authors requiring further information regarding Elsevier’s archiving and manuscript policies are encouraged to visit: http://www.elsevier.com/copyright Author's personal copy Cretaceous Research 32 (2011) 623e645 Contents lists available at ScienceDirect Cretaceous Research journal homepage: www.elsevier.com/locate/CretRes Cretaceous climate oscillations in the southern palaeolatitudes: New stable isotope evidence from India and Madagascar Yuri D. Zakharov a,*, Yasunari Shigeta b, Raghavendramurthy Nagendra c, Peter P. Safronov a, Olga P. Smyshlyaeva a, Alexander M. Popov a, Tatiana A. Velivetskaya a, Tamara B. Afanasyeva a a Far Eastern Geological Institute of Russian Academy of Sciences (Far Eastern Branch), Stoletiya Prospect 159, Vladivostok 690022, Russia b National Museum of Nature and Science, 3-23-1 Hyakunin-cho, Shinjuku-ku, Tokyo 169-0073, Japan c Department of Geology, Anna University, Chennai 600025, India article info abstract Article history: Palaeotemperatures for the Cretaceous of India and Madagascar have been determined on the basis of Received 18 March 2010 oxygen isotopic analysis of well-preserved Albian belemnite rostra and Maastrichtian bivalve shells of Accepted in revised form 18 April 2011 from the Trichinopoly district, southern India, and Albian nautiloid and ammonoid cephalopods from the Available online 23 April 2011 Mahajang Province, Madagascar. -

An Inventory of Belemnites Documented in Six Us National Parks in Alaska
Lucas, S. G., Hunt, A. P. & Lichtig, A. J., 2021, Fossil Record 7. New Mexico Museum of Natural History and Science Bulletin 82. 357 AN INVENTORY OF BELEMNITES DOCUMENTED IN SIX US NATIONAL PARKS IN ALASKA CYNTHIA D. SCHRAER1, DAVID J. SCHRAER2, JUSTIN S. TWEET3, ROBERT B. BLODGETT4, and VINCENT L. SANTUCCI5 15001 Country Club Lane, Anchorage AK 99516; -email: [email protected]; 25001 Country Club Lane, Anchorage AK 99516; -email: [email protected]; 3National Park Service, Geologic Resources Division, 1201 Eye Street, Washington, D.C. 20005; -email: justin_tweet@ nps.gov; 42821 Kingfisher Drive, Anchorage, AK 99502; -email: [email protected];5 National Park Service, Geologic Resources Division, 1849 “C” Street, Washington, D.C. 20240; -email: [email protected] Abstract—Belemnites (order Belemnitida) are an extinct group of coleoid cephalopods, known from the Jurassic and Cretaceous periods. We compiled detailed information on 252 occurrences of belemnites in six National Park Service (NPS) areas in Alaska. This information was based on published literature and maps, unpublished U.S. Geological Survey internal fossil reports (“Examination and Report on Referred Fossils” or E&Rs), the U.S. Geological Survey Mesozoic locality register, the Alaska Paleontological Database, the NPS Paleontology Archives and our own records of belemnites found in museum collections. Few specimens have been identified and many consist of fragments. However, even these suboptimal specimens provide evidence that belemnites are present in given formations and provide direction for future research. Two especially interesting avenues for research concern the time range of belemnites in Alaska. Belemnites are known to have originated in what is now Europe in the Early Jurassic Hettangian and to have a well-documented world-wide distribution in the Early Jurassic Toarcian. -

GY 112L Earth History
GY 112L Earth History Lab 11 The Mesozoic: Part Two GY 112L Instructors: Douglas Haywick, James Connors, Mary Anne Connors Department of Earth Sciences, University of South Alabama Fifth Edition: August 2009© The Fine Print Contents of these lab exercises are the intellectual property of the authors, particularly Dr. Doug Haywick. Contents cannot be reproduced outside of the University of South Alabama “family” (faculty and students) without the permission of D. Haywick. Internet users can seek this permission by contacting Dr. Haywick through the web address provided below. This manual is constantly being updated and occasionally, even improved. Typos, grammatical errors and sections that make no sense whatsoever may, or may not, be intentional. If you find an error, show it to your instructor. You may get bonus points. More likely you will be told to go away The recipes that are included in some sections are intended to prove that you can eat anything as long as you serve it with plenty of ketchup. Neither Haywick, nor the Connors are responsible for any food poisoning that might occur if you actually try them. http:/www.southalabama.edu/geology/haywick 1 Week Eleven The Mesozoic, Part Two: Molluscs (Cephalopods), Echinoderms and Alabama Stratigraphy Background: Well the good news is that we are rapidly approaching the end of this course. The bad news is that we are rapidly approaching the end of this course and we still have several important animal groups to consider. When we last visited the molluscs two weeks ago, we only had time to consider the gastropods and bivalves. -

Cephalopod Reproductive Strategies Derived from Embryonic Shell Size
Biol. Rev. (2017), pp. 000–000. 1 doi: 10.1111/brv.12341 Cephalopod embryonic shells as a tool to reconstruct reproductive strategies in extinct taxa Vladimir Laptikhovsky1,∗, Svetlana Nikolaeva2,3,4 and Mikhail Rogov5 1Fisheries Division, Cefas, Lowestoft, NR33 0HT, U.K. 2Department of Earth Sciences Natural History Museum, London, SW7 5BD, U.K. 3Laboratory of Molluscs Borissiak Paleontological Institute, Russian Academy of Sciences, Moscow, 117997, Russia 4Laboratory of Stratigraphy of Oil and Gas Bearing Reservoirs Kazan Federal University, Kazan, 420000, Russia 5Department of Stratigraphy Geological Institute, Russian Academy of Sciences, Moscow, 119017, Russia ABSTRACT An exhaustive study of existing data on the relationship between egg size and maximum size of embryonic shells in 42 species of extant cephalopods demonstrated that these values are approximately equal regardless of taxonomy and shell morphology. Egg size is also approximately equal to mantle length of hatchlings in 45 cephalopod species with rudimentary shells. Paired data on the size of the initial chamber versus embryonic shell in 235 species of Ammonoidea, 46 Bactritida, 13 Nautilida, 22 Orthocerida, 8 Tarphycerida, 4 Oncocerida, 1 Belemnoidea, 4 Sepiida and 1 Spirulida demonstrated that, although there is a positive relationship between these parameters in some taxa, initial chamber size cannot be used to predict egg size in extinct cephalopods; the size of the embryonic shell may be more appropriate for this task. The evolution of reproductive strategies in cephalopods in the geological past was marked by an increasing significance of small-egged taxa, as is also seen in simultaneously evolving fish taxa. Key words: embryonic shell, initial chamber, hatchling, egg size, Cephalopoda, Ammonoidea, reproductive strategy, Nautilida, Coleoidea. -

Doguzhaeva Etal 2014 Embryo
Embryonic shell structure of Early–Middle Jurassic belemnites, and its significance for belemnite expansion and diversification in the Jurassic LARISA A. DOGUZHAEVA, ROBERT WEIS, DOMINIQUE DELSATE AND NINO MARIOTTI Doguzhaeva, L.A., Weis, R., Delsate, D. & Mariotti N. 2014: Embryonic shell structure of Early–Middle Jurassic belemnites, and its significance for belemnite expansion and diversification in the Jurassic. Lethaia, Vol. 47, pp. 49–65. Early Jurassic belemnites are of particular interest to the study of the evolution of skel- etal morphology in Lower Carboniferous to the uppermost Cretaceous belemnoids, because they signal the beginning of a global Jurassic–Cretaceous expansion and diver- sification of belemnitids. We investigated potentially relevant, to this evolutionary pat- tern, shell features of Sinemurian–Bajocian Nannobelus, Parapassaloteuthis, Holcobelus and Pachybelemnopsis from the Paris Basin. Our analysis of morphological, ultrastruc- tural and chemical traits of the earliest ontogenetic stages of the shell suggests that modified embryonic shell structure of Early–Middle Jurassic belemnites was a factor in their expansion and colonization of the pelagic zone and resulted in remarkable diversification of belemnites. Innovative traits of the embryonic shell of Sinemurian– Bajocian belemnites include: (1) an inorganic–organic primordial rostrum encapsulating the protoconch and the phragmocone, its non-biomineralized compo- nent, possibly chitin, is herein detected for the first time; (2) an organic rich closing membrane which was under formation. It was yet perforated and possessed a foramen; and (3) an organic rich pro-ostracum earlier documented in an embryonic shell of Pliensbachian Passaloteuthis. The inorganic–organic primordial rostrum tightly coat- ing the protoconch and phragmocone supposedly enhanced protection, without increase in shell weight, of the Early Jurassic belemnites against explosion in deep- water environment. -

Abbreviation Kiel S. 2005, New and Little Known Gastropods from the Albian of the Mahajanga Basin, Northwestern Madagaskar
1 Reference (Explanations see mollusca-database.eu) Abbreviation Kiel S. 2005, New and little known gastropods from the Albian of the Mahajanga Basin, Northwestern Madagaskar. AF01 http://www.geowiss.uni-hamburg.de/i-geolo/Palaeontologie/ForschungImadagaskar.htm (11.03.2007, abstract) Bandel K. 2003, Cretaceous volutid Neogastropoda from the Western Desert of Egypt and their place within the noegastropoda AF02 (Mollusca). Mitt. Geol.-Paläont. Inst. Univ. Hamburg, Heft 87, p 73-98, 49 figs., Hamburg (abstract). www.geowiss.uni-hamburg.de/i-geolo/Palaeontologie/Forschung/publications.htm (29.10.2007) Kiel S. & Bandel K. 2003, New taxonomic data for the gastropod fauna of the Uzamba Formation (Santonian-Campanian, South AF03 Africa) based on newly collected material. Cretaceous research 24, p. 449-475, 10 figs., Elsevier (abstract). www.geowiss.uni-hamburg.de/i-geolo/Palaeontologie/Forschung/publications.htm (29.10.2007) Emberton K.C. 2002, Owengriffithsius , a new genus of cyclophorid land snails endemic to northern Madagascar. The Veliger 45 (3) : AF04 203-217. http://www.theveliger.org/index.html Emberton K.C. 2002, Ankoravaratra , a new genus of landsnails endemic to northern Madagascar (Cyclophoroidea: Maizaniidae?). AF05 The Veliger 45 (4) : 278-289. http://www.theveliger.org/volume45(4).html Blaison & Bourquin 1966, Révision des "Collotia sensu lato": un nouveau sous-genre "Tintanticeras". Ann. sci. univ. Besancon, 3ème AF06 série, geologie. fasc.2 :69-77 (Abstract). www.fossile.org/pages-web/bibliographie_consacree_au_ammon.htp (20.7.2005) Bensalah M., Adaci M., Mahboubi M. & Kazi-Tani O., 2005, Les sediments continentaux d'age tertiaire dans les Hautes Plaines AF07 Oranaises et le Tell Tlemcenien (Algerie occidentale). -

The Gladiuses in Coleoid Cephalopods: Homology, Parallelism, Or Convergence?
Swiss J Palaeontol (2015) 134:187–197 DOI 10.1007/s13358-015-0100-3 The gladiuses in coleoid cephalopods: homology, parallelism, or convergence? 1 1 Dirk Fuchs • Yasuhiro Iba Received: 26 February 2015 / Accepted: 15 August 2015 / Published online: 25 September 2015 Ó Akademie der Naturwissenschaften Schweiz (SCNAT) 2015 Abstract In the cephalopod subclass Coleoidea, several cuttlebone demonstrates furthermore that a gladius might homology problems exist, mainly owing to unsolved phy- have also evolved from a secondarily proostracum-less logenetic relationships between decabrachian orders. The phragmocone. Life styles and habitats of living and present contribution reviews the ‘‘similarity’’ of the glad- Mesozoic gladius-bearing octobrachians are finally dis- ius, the chitinous shell rudiment in the dorsal mantle that cussed in the light of our conclusions. provides rigid attachment sites for the locomotory-relevant musculature. As a secretion product of the shell sac Keywords Cephalopoda Á Coleoidea Á Gladiuses Á epithelium as well as in the light of a common three-lay- Homology Á Parallelism Á Convergence ered construction, both the octobrachian and the deca- brachian gladius types most probably represent homologues with identical developmental mechanisms; Introduction ‘‘similarities’’ in gladius shapes in unrelated lineages therefore should be considered as the result of parallelism. The gladius (English ‘‘pen’’; French ‘‘plume’’; German Ultrastructural comparisons with Mesozoic coleoids sug- ‘‘Schulp’’) is a chitinous, spatulate structure located in the gest that an organic gladius is actually embedded in every dorsal midline of the body of coleoid cephalopods. This proostracum-bearing phragmocone. It is therefore gener- sturdy ‘‘backbone’’ typically occupies the full length of the ally accepted that a gladius evolved through decalcification dorsal mantle and provides attachment of various loco- of a proostracum-bearing phragmocone. -

Published Version
[Palaeontology, Vol. 54, Part 2, 2011, pp. 397–415] THE CAPSULE: AN ORGANIC SKELETAL STRUCTURE IN THE LATE CRETACEOUS BELEMNITE GONIOTEUTHIS FROM NORTH-WEST GERMANY by LARISA A. DOGUZHAEVA and STEFAN BENGTSON Department of Palaeozoology, Swedish Museum of Natural History, PO Box 50007, SE-104 05 Stockholm, Sweden; e-mails [email protected]; [email protected] Typescript received 7 February 2009; accepted in revised form 1 June 2010 Abstract: An unusual, bilaterally symmetrical black struc- A flare along the periphery of the alveolus marks a region ture that embraces the protoconch and the phragmocone where the rostrum was not yet formed, suggesting that the and is overlain by a rostrum has been studied in the Santo- capsule extended beyond the rostrum. Modification of the nian–early Campanian (Late Cretaceous) belemnite genus skeleton in Gonioteuthis comprises a set of supposedly inter- Gonioteuthis from Braunschweig, north-west Germany. The related changes, such as innovation of the organic capsule, structure is here named the capsule. Energy dispersed spec- partial elimination of the calcareous rostrum and a diminish- trometry analyses of the capsule show a co-occurrence of sul- ing of the pro-ostracum, resulting in the appearance of a phur with zinc, barium, iron, lead and titanium, suggesting new type of pro-ostracum that became narrower and shorter their chemical association. The capsule was originally made and lost the spatula-like shape and gently curved growth of organic material that was diagenetically transformed into lines of a median field that are typical for the majority of sulphur-containing matter. The material of the capsule dif- Jurassic and Cretaceous belemnites. -

Maastrichtian Ceratisepia and Mesozoic Cuttlebone Homeomorphs
Maastrichtian Ceratisepia and Mesozoic cuttlebone homeomorphs ROGER A. HEWITT and JOHN W.M. JAGT Hewitt, R.A. & Jagt, J.W.M. 1999. Maastrichtian Ceratisepia and Mesozoic cuttlebone homeomorphs. -Acta Palaeontologica Polonica 44,3,305-326. The phylogenetics of potential Mesozoic ancestors of cuttlefish of a restricted order Sepiida von Zittel, 1895 (superorder Decabrachia Boettger, 1952) is reviewed. Micro- structural studies of Mesozoic homeomorphs of cuttlebones (Pearceiteuthis gen. n., Loligosepia, Trachyteuthis, Actinosepia) are consistent with their assignement to the superorder Octobrachia Fioroni, 1981. The discovery of an embryonic Ceratisepia shell in the upper Maastrichtian of the Netherlands, indicates that true Sepiida did have a pre-Cenozoic origin. Cretaceous decabrachs of the order Spirulida Stolley, 1919 do not show evidence of the dorso-anterior shell growth vectors seen in Cenozoic spirulids, sepiids and octobrachs. Separate origins of the Sepiida and Spirulida within Cretaceous diplobelinid belemnites is still the most attractive hypothesis. Ceratisepia vanknippen- bergi sp. n. from the upper Maastrichtian of the Netherlands and Pearceiteuthis buyi gen. et sp. n. from the Callovian of England are described. Key words : Cephalopods, cuttlefish, ontogeny, biornineralization, Mesozoic. Roger A. Hewitt, 12 Fairj-leld Road, Eastwood, Leigh-on-Sea, Essex SS9 5SB, United Kingdom. John W.M. Jagt [[email protected]], Natuurhistorisch Museum Maastricht, P.O. Box 882, NL-6200 AW Maastricht, the Netherlands. Introduction Cuttlebones are those dorsally hard and spherulitic, but ventrally soft and septate, aragonitic shells that invade beaches (Cadee 1997) and can be seen being sold to the owners of caged birds in pet shops. They are the expanded and highly modified inter- nal phragmocones of cephalopods (Bandel & von Boletzky 1979) and belong to a di- verse group of cuttlefish that appeared in the Rupelian (Szordnyi 1934; Hewitt & Pedley 1978; Engeser 1990). -

Late Cretaceous Record of Large Soft-Bodied Coleoids Based on Lower Jaw Remains from Hokkaido, Japan
Editors' choice Late Cretaceous record of large soft-bodied coleoids based on lower jaw remains from Hokkaido, Japan KAZUSHIGE TANABE, AKIHIRO MISAKI, and TAKAO UBUKATA Tanabe, K., Misaki, A., and Ubukata, T. 2015. Late Cretaceous record of large soft-bodied coleoids based on lower jaw remains from Hokkaido, Japan. Acta Palaeontologica Polonica 60 (1): 27–38. The origin and phylogenetic relationships of most modern coleoid groups have not yet been explained by reliable fossil evidence, in large part because of the reduction or disappearance of a calcified chambered shell during their evolutionary history. Herein we describe two exceptionally large coleoid lower jaws from the Upper Cretaceous strata in Hokkaido, Japan. On the basis of the comparison of gross morphology and morphometric data of the lower jaws of modern and fossil coleoids, we assigned the two lower jaws to the following new taxa: Nanaimoteuthis hikidai sp. nov. of the order Vampyromorpha (superorder Octobrachia) and Haboroteuthis poseidon gen. et sp. nov. of the order Teuthida (super- order Decabrachia). The lower jaw of N. hikidai is distinguished from other species of the same genus from the Upper Cretaceous of Vancouver Island (Canada) and Hokkaido by having a broader, more anteriorly curved hood of the outer lamella. The lower jaw of H. poseidon seemingly exhibits mosaic features like those of modern teuthids and sepiids but is assigned to Teuthida on the basis of the overall shape of the outer lamella and the development of a distinct fold on the lateral wall. Because of the unusually large lower jaws, these new taxa appear to be comparable in body size to modern giant squids (Architeuthis spp.) and the Humboldt squid (Dosidicus gigas). -

Cephalopod Classification and Taxonomy
Cephalopod classification and Taxonomy 09 V. Venkatesan and K.S. Mohamed Molluscan Fisheries Division, Central Marine Fisheries Research Institute, Kochi-682 018 Introduction have proved to be useful based on the morphological studies Chambered nautilus, cuttlefishes, squids and octopus are (Voss, 1988), but less significant in determining higher-level the four major groups of cephalopods, which belong to the relationships. The following classification can be used until an highly evolved class of phylum Mollusca. Cephalopods are the alternative arrangement can be derived from cladistic analysis. third largest molluscan class after bivalves and gastropods Molecular or morphological based analysis must have to go and consist of more than 800 species (Lindgren et al. 2004). considerable scrutiny before changed in classification should The fossil record contains about 17,000 named species of be adopted. cephalopods. Although the diversity of cephalopods is very much reduced in the modern era, cephalopods are found to Cephalopods (Class Cephalopoda) are represented by two occur in all the oceans of the world from the tropics to the extant subclasses, Nautiloidea (Nautilus and Allonautilus) and polar seas and at all depths ranging from the surface to below Coleoidea and one extinct subclass, Ammonoidea. Members 5000m. Cephalopods were dominant predators millions of the subclass Coleoidea includes two subdivisions, the of years before fish appeared. The earliest cephalopods Belemnoidea, which is the primitive form of cephalopod were primitive shelled nautiloids which evolved in the Late possessing ink sac and ten equally sized arms, became extinct Cambrian period. The living cephalopods range in size from during the cretaceous period and Neocoleoidea (cuttlefish, 25mm (Southern pygmy squid, Idiosepius notoides) to more squid and octopus) where the shell has been internalized than 12m (Colossal squid, Mesonychoteuthis hamiltoni) in and reduced, completely lost. -
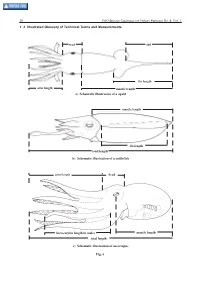
1.4 Illustrated Glossary of Technical Terms and Measurements
20 FAO Species Catalogue for Fishery Purposes No. 4, Vol. 1 1. 4 Illustrated Glossary of Technical Terms and Measurements head tail fin length arm length mantle length a) Schematic illustration of a squid mantle length fin length total length b) Schematic illustration of a cuttlefish arm length head hectocotylus length in males mantle length total length c) Schematic illustration of an octopus Fig. 6 Cephalopods of the World 21 Abdominal septum – Median septum that traverses the Apomorphic – Derived from a more ancestral condition. mantle cavity parallel to the body axis. It extends from the Loosely considered the ‘advanced’ condition. visceral mass to the ventral mantle wall. The ventral mantle artery runs along the dorsal edge of this septum. Arm – One of the circumoral appendages of cephalopods. Arms are designated by the numbers I to IV, starting with I Aboral – Away from or opposite to the mouth. as the dorsal (or upper) pair. In decapodiformes each appendage of the fourth original pair is modified to form a Abyssal – The great depths of the ocean: from 2 000 to tentacle. The second pair of arms is modified in vampires 6 000 m. into a long filamentous structure. It has been lost in cirrate and incirrate octopods. Accessory nidamental glands – Glands of unknown function; consist of tubules containing symbiotic bacteria. Arm formula – Comparative length of the 4 pairs of arms Found in all decapodiformes except oegopsid squids. expressed numerically in decreasing order: the largest arm is indicated first and the shortest last, e.g. IV>III>II>I. If Adult – A female that has mature eggs (these frequently are IV>III=II>I, then arm IV is the longest, followed by arm III stored in the oviducts), or a male that has produced which is the same size as arm II and both are longer than spermatophores (these are stored in Needham’s sac).