Larval Fish Dispersal and the East Pacific Barrier
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Pacific Plate Biogeography, with Special Reference to Shorefishes
Pacific Plate Biogeography, with Special Reference to Shorefishes VICTOR G. SPRINGER m SMITHSONIAN CONTRIBUTIONS TO ZOOLOGY • NUMBER 367 SERIES PUBLICATIONS OF THE SMITHSONIAN INSTITUTION Emphasis upon publication as a means of "diffusing knowledge" was expressed by the first Secretary of the Smithsonian. In his formal plan for the Institution, Joseph Henry outlined a program that included the following statement: "It is proposed to publish a series of reports, giving an account of the new discoveries in science, and of the changes made from year to year in all branches of knowledge." This theme of basic research has been adhered to through the years by thousands of titles issued in series publications under the Smithsonian imprint, commencing with Smithsonian Contributions to Knowledge in 1848 and continuing with the following active series: Smithsonian Contributions to Anthropology Smithsonian Contributions to Astrophysics Smithsonian Contributions to Botany Smithsonian Contributions to the Earth Sciences Smithsonian Contributions to the Marine Sciences Smithsonian Contributions to Paleobiology Smithsonian Contributions to Zoo/ogy Smithsonian Studies in Air and Space Smithsonian Studies in History and Technology In these series, the Institution publishes small papers and full-scale monographs that report the research and collections of its various museums and bureaux or of professional colleagues in the world cf science and scholarship. The publications are distributed by mailing lists to libraries, universities, and similar institutions throughout the world. Papers or monographs submitted for series publication are received by the Smithsonian Institution Press, subject to its own review for format and style, only through departments of the various Smithsonian museums or bureaux, where the manuscripts are given substantive review. -

Updated Checklist of Marine Fishes (Chordata: Craniata) from Portugal and the Proposed Extension of the Portuguese Continental Shelf
European Journal of Taxonomy 73: 1-73 ISSN 2118-9773 http://dx.doi.org/10.5852/ejt.2014.73 www.europeanjournaloftaxonomy.eu 2014 · Carneiro M. et al. This work is licensed under a Creative Commons Attribution 3.0 License. Monograph urn:lsid:zoobank.org:pub:9A5F217D-8E7B-448A-9CAB-2CCC9CC6F857 Updated checklist of marine fishes (Chordata: Craniata) from Portugal and the proposed extension of the Portuguese continental shelf Miguel CARNEIRO1,5, Rogélia MARTINS2,6, Monica LANDI*,3,7 & Filipe O. COSTA4,8 1,2 DIV-RP (Modelling and Management Fishery Resources Division), Instituto Português do Mar e da Atmosfera, Av. Brasilia 1449-006 Lisboa, Portugal. E-mail: [email protected], [email protected] 3,4 CBMA (Centre of Molecular and Environmental Biology), Department of Biology, University of Minho, Campus de Gualtar, 4710-057 Braga, Portugal. E-mail: [email protected], [email protected] * corresponding author: [email protected] 5 urn:lsid:zoobank.org:author:90A98A50-327E-4648-9DCE-75709C7A2472 6 urn:lsid:zoobank.org:author:1EB6DE00-9E91-407C-B7C4-34F31F29FD88 7 urn:lsid:zoobank.org:author:6D3AC760-77F2-4CFA-B5C7-665CB07F4CEB 8 urn:lsid:zoobank.org:author:48E53CF3-71C8-403C-BECD-10B20B3C15B4 Abstract. The study of the Portuguese marine ichthyofauna has a long historical tradition, rooted back in the 18th Century. Here we present an annotated checklist of the marine fishes from Portuguese waters, including the area encompassed by the proposed extension of the Portuguese continental shelf and the Economic Exclusive Zone (EEZ). The list is based on historical literature records and taxon occurrence data obtained from natural history collections, together with new revisions and occurrences. -

Atoll Research Bulletin No. 548
ATOLL RESEARCH BULLETIN NO. 548 STOMACH CONTENTS AND FEEDING OBSERVATIONS OF SOME EASTER ISLAND FISHES BY LOUIS H. DISALVO, JOHN E. RANDALL, AND ALFREDO CEA ISSUED BY NATIONAL MUSEUM OF NATURAL HISTORY SMITHSONIAN INSTITUTION WASHINGTON, D.C. U.S.A. DECEMBER 2007 STOMACH CONTENTS AND FEEDING OBSERVATIONS OF SOME EASTER ISLAND FISHES BY LOUIS H. DISALVO,1 JOHN E. RANDALL,2 AND ALFREDO CEA3 ABSTRACT Stomach contents of 42 species in 25 familes of Easter Island shore fishes were examined in comparative terms to determine prey items and feeding behavior at this isolated island outpost in the southeastern Pacific. The island’s impoverished marine fauna and flora have resulted in considerable dietary overlap among the inshore fishes. Some endemic species appear to feed mainly on endemic invertebrates. Some prey species which were found in the fish stomachs, such as the stomatopodOdontodactylus hawaiiensis, the pandalid shrimp Plesionika edwardsi, and several tiny molluscs were previously unrecorded for the island. INTRODUCTION Easter Island (Rapa Nui) lies 3750 km west of the South American continent and 2250 km. East of the Pitcairn Islands. This island represents the most isolated landfall in the South Pacific Ocean along with its small rocky neighbor Salas y Gómez I. 415 km to the east. Although Easter Island is often regarded as part of the Indo-Pacific region, and most of its fauna consist of tropical species (in the case of shore fishes, 32.5%), it lies outside the 20° isotherm (Wells 1957) where seawater temperature and insolation are below that required for the development of structural coral reefs; water temperature can undergo interannual drops unfavorable to tropical organisms and may produce mass mortalities of corals (Wellington et al., 2001). -

Annotated Checklist of the Fish Species (Pisces) of La Réunion, Including a Red List of Threatened and Declining Species
Stuttgarter Beiträge zur Naturkunde A, Neue Serie 2: 1–168; Stuttgart, 30.IV.2009. 1 Annotated checklist of the fish species (Pisces) of La Réunion, including a Red List of threatened and declining species RONALD FR ICKE , THIE rr Y MULOCHAU , PA tr ICK DU R VILLE , PASCALE CHABANE T , Emm ANUEL TESSIE R & YVES LE T OU R NEU R Abstract An annotated checklist of the fish species of La Réunion (southwestern Indian Ocean) comprises a total of 984 species in 164 families (including 16 species which are not native). 65 species (plus 16 introduced) occur in fresh- water, with the Gobiidae as the largest freshwater fish family. 165 species (plus 16 introduced) live in transitional waters. In marine habitats, 965 species (plus two introduced) are found, with the Labridae, Serranidae and Gobiidae being the largest families; 56.7 % of these species live in shallow coral reefs, 33.7 % inside the fringing reef, 28.0 % in shallow rocky reefs, 16.8 % on sand bottoms, 14.0 % in deep reefs, 11.9 % on the reef flat, and 11.1 % in estuaries. 63 species are first records for Réunion. Zoogeographically, 65 % of the fish fauna have a widespread Indo-Pacific distribution, while only 2.6 % are Mascarene endemics, and 0.7 % Réunion endemics. The classification of the following species is changed in the present paper: Anguilla labiata (Peters, 1852) [pre- viously A. bengalensis labiata]; Microphis millepunctatus (Kaup, 1856) [previously M. brachyurus millepunctatus]; Epinephelus oceanicus (Lacepède, 1802) [previously E. fasciatus (non Forsskål in Niebuhr, 1775)]; Ostorhinchus fasciatus (White, 1790) [previously Apogon fasciatus]; Mulloidichthys auriflamma (Forsskål in Niebuhr, 1775) [previously Mulloidichthys vanicolensis (non Valenciennes in Cuvier & Valenciennes, 1831)]; Stegastes luteobrun- neus (Smith, 1960) [previously S. -

Chec List Marine and Coastal Biodiversity of Oaxaca, Mexico
Check List 9(2): 329–390, 2013 © 2013 Check List and Authors Chec List ISSN 1809-127X (available at www.checklist.org.br) Journal of species lists and distribution ǡ PECIES * S ǤǦ ǡÀ ÀǦǡ Ǧ ǡ OF ×±×Ǧ±ǡ ÀǦǡ Ǧ ǡ ISTS María Torres-Huerta, Alberto Montoya-Márquez and Norma A. Barrientos-Luján L ǡ ǡǡǡǤͶǡͲͻͲʹǡǡ ǡ ȗ ǤǦǣ[email protected] ćĘęėĆĈęǣ ϐ Ǣ ǡǡ ϐǤǡ ǤǣͳȌ ǢʹȌ Ǥͳͻͺ ǯϐ ʹǡͳͷ ǡͳͷ ȋǡȌǤǡϐ ǡ Ǥǡϐ Ǣ ǡʹͶʹȋͳͳǤʹΨȌ ǡ groups (annelids, crustaceans and mollusks) represent about 44.0% (949 species) of all species recorded, while the ʹ ȋ͵ͷǤ͵ΨȌǤǡ not yet been recorded on the Oaxaca coast, including some platyhelminthes, rotifers, nematodes, oligochaetes, sipunculids, echiurans, tardigrades, pycnogonids, some crustaceans, brachiopods, chaetognaths, ascidians and cephalochordates. The ϐϐǢ Ǥ ēęėĔĉĚĈęĎĔē Madrigal and Andreu-Sánchez 2010; Jarquín-González The state of Oaxaca in southern Mexico (Figure 1) is and García-Madrigal 2010), mollusks (Rodríguez-Palacios known to harbor the highest continental faunistic and et al. 1988; Holguín-Quiñones and González-Pedraza ϐ ȋ Ǧ± et al. 1989; de León-Herrera 2000; Ramírez-González and ʹͲͲͶȌǤ Ǧ Barrientos-Luján 2007; Zamorano et al. 2008, 2010; Ríos- ǡ Jara et al. 2009; Reyes-Gómez et al. 2010), echinoderms (Benítez-Villalobos 2001; Zamorano et al. 2006; Benítez- ϐ Villalobos et alǤʹͲͲͺȌǡϐȋͳͻͻǢǦ Ǥ ǡ 1982; Tapia-García et alǤ ͳͻͻͷǢ ͳͻͻͺǢ Ǧ ϐ (cf. García-Mendoza et al. 2004). ǡ ǡ studies among taxonomic groups are not homogeneous: longer than others. Some of the main taxonomic groups ȋ ÀʹͲͲʹǢǦʹͲͲ͵ǢǦet al. -

Peces Óseos Marinos (Pisces: Osteichthyes)
Bibliografía sobre Biodiversidad Acuática de Chile (Pisces: Os P eces ÓseosMarinos t eich th y es) Bibliografía sobre Biodiversidad Acuática de Chile Palma, S., P. Báez & G. Pequeño (eds.). 2010 Comité Oceanográfico Nacional, Valparaíso, pp. 315-395 Peces Óseos Marinos (Pisces: Osteichthyes) Germán Pequeño1, Sylvia Sáez1 & Gloria Pizarro2 1Instituto de Zoología “Ernst F. Kilian”, Universidad Austral de Chile. Casilla 567, Valdivia, E-mail: [email protected] 2Librería Punto 4, Valdivia. RESUMEN Se presentan 1.772 referencias sobre taxonomía, sistemática y distribución geográfica de los peces óseos marinos y que siendo marinos penetran en estuarios de Chile. Se consideran peces de la costa de Chile en el continente sudamericano y también de las islas oceánicas chilenas. Palabras clave: peces, bibliografía, biodiversidad, Pacífico suroriental, Chile. ABSTRACT One thousand seven hundred and seventy-two references on taxonomy, systematics and geographical distribution of marine bony fishes and those that being marine enter in estuaries of Chile are presented. The species considered are those from the Chilean continental coast and those from the Chilean oceanic islands. Key words: fishes, bibliography, biodiversity, southeastern, Chile. INTRODUCCIÓN marinos, incursionan en estuarios, consi-derando la costa de Chile que integra el continente sudamericano Actualmente, se podría decir que las especies y aquella de las islas oceánicas. Puede observarse conocidas de peces óseos de aguas marinas chilenas que se trata de un listado bastante restringido, pero alcanzan un número cercano a las 1.050. Se supone que servirá al lector, en los aspectos científicos que dada la falta de exploración, especialmente en el básicos señalados. dominio marino, ese número debería aumentar. -
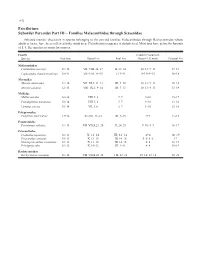
Perciformes Suborder Percoidei Part III – Families Malacanthidae
1152 Perciformes Suborder Percoidei Part III – Families Malacanthidae through Sciaenidae Selected meristic characters in species belonging to the percoid families Malacanthidae through Rachycentridae whose adults or larvae have been collected in the study area. Classification sequence is alphabetical. Most taxa have pelvic fin formula of I, 5. See species accounts for sources. Family Caudal (Procurrent, Species Vertebrae Dorsal Fin Anal Fin Dorsal + Ventral) Pectoral Fin Malacanthidae Caulolatilus microps 11+16 VII–VIII, 24–27 II, 22–24 10–13+9–13 17–18 Lopholatilus chamaeleonticeps 10+14 VII–VIII, 14–15 I, 13–14 9–13+9–13 16–18 Moronidae Morone americana 11+14 VII–XI, I, 11–13 III, 9–10 10–13+9–13 10–18 Morone saxatilis 12+13 VIII–IX, I, 9–14 III, 7–13 10–13+9–13 13–19 Mullidae Mullus auratus 10+14 VIII, I, 8 I, 7 9+10 15–17 Pseudupeneus maculatus 10+14 VIII, I, 8 I, 7 9+10 13–16 Upeneus parvus 10+14 VII, I, 8 I, 7 9+10 15–16 Polyprionidae Polyprion americanus 13+14 XI–XII, 11–12 III, 9–10 9+9 17–18 Pomatomidae Pomatomus saltatrix 11+15 VII–VIII,I,23–28 II, 24–29 9–10+8–9 16–17 Priacanthidae Cookeolus japonicus 10+13 X, 12–14 III, 12–14 4+4 18–19 Priacanthus arenatus 10+13 X, 13–15 III, 14–16 5–6+5–6 17 Heteropriacanthus cruentatus 10+13 X, 13–14 III, 14–15 4+4 18–19 Pristigenys alta 10+13 X, 10–12 III, 9–11 4+4 16–19 Rachycentridae Rachycentron canadum 11+14 VII–VIII,I,26–34 I–II, 22–28 15–16+12–14 20–21 Early Stages of Fishes in the Western North Atlantic Ocean 1153 Perciformes Suborder Percoidei Part III – Families Malacanthidae through Sciaenidae Selected meristic characters in species belonging to the percoid family Sciaenidae whose adults or larvae have been col- lected in the study area. -

Check List LISTS of SPECIES Check List 11(3): 1659, May 2015 Doi: ISSN 1809-127X © 2015 Check List and Authors
11 3 1659 the journal of biodiversity data May 2015 Check List LISTS OF SPECIES Check List 11(3): 1659, May 2015 doi: http://dx.doi.org/10.15560/11.3.1659 ISSN 1809-127X © 2015 Check List and Authors Coastal fishes of Rio Grande do Norte, northeastern Brazil, with new records José Garcia Júnior1*, Marcelo Francisco Nóbrega2 and Jorge Eduardo Lins Oliveira2 1 Instituto Federal de Educação, Ciência e Tecnologia do Rio Grande do Norte, Campus Macau, Rua das Margaridas, 300, CEP 59500-000, Macau, RN, Brazil 2 Universidade Federal do Rio Grande do Norte, Departamento de Oceanografia e Limnologia, Laboratório de Biologia Pesqueira, Praia de Mãe Luiza, s/n°, CEP 59014-100, Natal, RN, Brazil * Corresponding author. E-mail: [email protected] Abstract: An updated and reviewed checklist of coastal more continuous than northern coast, the three major fishes of the Rio Grande do Norte state, northeastern estuaries are small and without many ramifications, and coast of Brazil, is presented. Between 2003 and 2013 the reefs are more numerous but smaller and relatively the occurrence of fish species were recorded through closer to each other than northern coast. The first and collection of specimens, landing records of the artisanal only checklist of fish species that occur along the coast fleet, literature reviews and from specimens deposited of RN was produced in 1988 and comprised 190 species in ichthyological collections. A total of 459 species from (Soares 1988). This situation improved after 2000 with 2 classes, 26 orders, 102 families and 264 genera is listed, fish surveys in specific sites of the coast (e.g., Feitoza with 83 species (18% of the total number) recorded for 2001; Feitosa et al. -

Revision of the Circumtropical Glasseye Fish Heteropriacanthus Cruentatus (Perciformes: Priacanthidae), with Resurrection of Two Species
Zootaxa 4273 (3): 341–361 ISSN 1175-5326 (print edition) http://www.mapress.com/j/zt/ Article ZOOTAXA Copyright © 2017 Magnolia Press ISSN 1175-5334 (online edition) https://doi.org/10.11646/zootaxa.4273.3.2 http://zoobank.org/urn:lsid:zoobank.org:pub:9C71FA99-2117-4E56-89DD-5E6B6FB2894E Revision of the circumtropical glasseye fish Heteropriacanthus cruentatus (Perciformes: Priacanthidae), with resurrection of two species IRIA FERNANDEZ-SILVA1,2,3 & HSUAN-CHING HO3,4* 1California Academy of Sciences, San Francisco, U.S.A. 2Department of Biochemistry, Genetics and Immunology, University of Vigo, Spain 3National Museum of Marine Biology & Aquarium, Pingung, Taiwan 4Institute of Marine Biology, National Museum of Marine Biology, Pingtung, Taiwan 5Corresponding author. E-mail: [email protected] Abstract The glasseye fish Heteropriacanthus, previously known as a monotypic genus, is now divided into three species based on morphological and genetic features. After examination on the type specimens and literature, herein we resurrect two junior binomens, H. carolinus (Cuvier, 1829) from the Indo-Pacific Ocean and H. fulgens (Lowe, 1838) from the northeastern Atlantic Ocean. Heteropriacanthus cruentatus (Lacepède, 1801) is now considered to be restricted to the Atlantic and southwestern Indian oceans. In light of these observations we discuss the evolutionary history of the genus. Key words: Circumtropical, genetics, Heteropriacanthus carolinus, Heteropriacanthus fulgens, teleostei, taxonomy Inroduction The bigeye family Priacanthidae is a small group of small to mid-sized generally epibenthic fishes that occur in tropical and subtropical seas near coral reefs or rock formations at depths 5 to 400 m or deeper (Starnes, 1988). Members of this family are known to produce sound (Salmon & Winn, 1966). -

Tropical Transpacific Shore Fishesl
Tropical Transpacific Shore Fishesl D. Ross Robertson, 2 Jack S. Grove, 3 and John E. McCosker4 Abstract: Tropical transpacific fishes occur on both sides of the world's largest deep-water barrier to the migration of marine shore organisms, the 4,000- to 7,000-km-wide Eastern Pacific Barrier (EPB). They include 64 epipelagic oce anic species and 126 species ofshore fishes known from both the tropical eastern Pacific (TEP) and the central and West Pacific. The broad distributions of 19 of 39 circumglobal transpacific species ofshore fishes offer no clues to the origin of their TEP populations; TEP populations of another 19 with disjunct Pacific distributions may represent isthmian relicts that originated from New World populations separated by the closure of the Central American isthmus. Eighty species of transpacific shore fishes likely migrated eastward to the TEP, and 22 species of shore fishes (12 of them isthmian relicts) and one oceanic species likely migrated westward from the TEP. Transpacific species constitute ~12% of the TEP's tropical shore fishes and 15-20% of shore fishes at islands on the western edge of the EPB. Eastward migrants constitute ~ 7% of the TEP's shore-fish fauna, and a similar proportion of TEP endemics may be derived from recent eastward immigration. Representation of transpacific species in different elements of the TEP fauna relates strongly to adult pelagic dispersal ability-they constitute almost all the epipelagic oceanic species, ~25% of the inshore pelagic species, but only 10% of the demersal shore fishes. Taxa that have multiple pelagic life-history stages are best represented among the transpacific species. -

The Intermuscular Bones and Ligaments of Teleostean Fishes *
* The Intermuscular Bones and Ligaments of Teleostean Fishes COLIN PATTERSON and G. DAVID JOHNSON m I I SMITHSONIAN CONTRIBUTIONS TO ZOOLOGY • NUMBER 559 SERIES PUBLICATIONS OF THE SMITHSONIAN INSTITUTION Emphasis upon publication as a means of "diffusing knowledge" was expressed by the first Secretary of the Smithsonian. In his formal plan for the institution, Joseph Henry outlined a program that included the following statement: "It is proposed to publish a series of reports, giving an account of the new discoveries in science, and of the changes made from year to year in all branches of knowledge." This theme of basic research has been adhered to through the years by thousands of titles issued in series publications under the Smithsonian imprint, commencing with Smithsonian Contributions to Knowledge in 1848 and continuing with the following active series: Smithsonian Contributions to Anthropology Smithsonian Contributions to Botany Smithsonian Contributions to the Earth Sciences Smithsonian Contributions to the Marine Sciences Smithsonian Contributions to Paleobiology Smithsonian Contributions to Zoology Smithsonian Folklife Studies Smithsonian Studies in Air and Space Smithsonian Studies in History and Technology In these series, the Institution publishes small papers and full-scale monographs that report the research and collections of its various museums and bureaux or of professional colleagues in the world of science and scholarship. The publications are distributed by mailing lists to libraries, universities, and similar institutions throughout the world. Papers or monographs submitted for series publication are received by the Smithsonian Institution Press, subject to its own review for format and style, only through departments of the various Smithsonian museums or bureaux, where the manuscripts are given substantive review. -

South Atlantic Mpas and Oculina HAPC: Characterization of Fish Communities, Benthic Habitat, and Benthic Macrobiota
South Atlantic MPAs and Oculina HAPC: Characterization of Fish Communities, Benthic Habitat, and Benthic Macrobiota Final Report for 2015-2017 NOAA Ship Pisces Cruises (15-02, 16-20, 17-02) Funding: NOAA Coral Reef Conservation Program (CRCP) South Atlantic Fishery Management Council (SAFMC) NOAA CRCP-Fishery Management Council Coral Reef Conservation Cooperative Agreements Project ID#: NA14NMF4410149 Stacey Harter, Principal Investigator NMFS/Southeast Fisheries Science Center (SEFSC) Panama City, FL Email: [email protected] John Reed and Stephanie Farrington Harbor Branch Oceanographic Institute, Florida Atlantic University Fort Pierce, FL Email: [email protected] Andrew David NMFS/Southeast Fisheries Science Center (SEFSC) Panama City, FL Email: [email protected] Citation: Harter, Stacey, John Reed, Stephanie Farrington, Andy David. 2018. South Atlantic MPAs and Oculina HAPC: Characterization of fish communities, benthic habitat, and benthic marcrobiota. Final Report for 2015-2017 NOAA Ship Pisces Cruises (15-02, 16-20, 17-02). NOAA CIOERT Cruise Report, 97 pp. Harbor Branch Oceanographic Technical Report Number 186. June 27, 2018 TABLE OF CONTENTS Executive Summary ........................................................................................................................ 4 Acknowldegements ......................................................................................................................... 4 Deliverables and Data Management ..............................................................................................