Rainbow Peacock Spiders Inspire Miniature Super-Iridescent Optics
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Diffractive Properties of Blue Morpho Butterfly Wings
Diffractive Properties of Blue Morpho Butterfly Wings Mary Lalak and Paul Brackman Department of Physics and Astronomy, University of Georgia, Athens, Georgia 30602 (Dated: October 24, 2014) Many species of butterflies are known to produce beautiful iridescent colors when exposed to light from different angles. These affects can be attributed to a few different optical phenomena combined, the most prominent being reflective diffraction. Using common household items and with a budget of 50 dollars, we attempt to confirm the theory that the collection of ridges on the individual scales on the wing act as transmission gratings, as well as reflective. These results are quantified by the calculation of the ridge separation and comparing this distance to those observed by a scanning electron microscope photo. I. INTRODUCTION In studying the optical property of iridescence, three distinct mechanisms must be discussed. Thin film inter- ference, structured coloring, and reflective diffraction all contribute to the iridescent qualities of a surface. Thin film interference occurs when light strikes a film, some of it enters the film, and the rest is reflected off. The light transmitted through reflects off the bottom of the film and exits the film to interfere with the light that FIG. 1: Up-Close View of Scales from an Optical Microscope originally reflected off the surface of the film. This inter- ference pattern causes a spectrum of color to be visible from white light. Examples of thin film interference in- clude oil slicks and soap bubbles. Structural coloring oc- curs when the structure of the object itself (with various reflective surfaces) produces an interference resulting in vibrant colors. -

Iridescence in Cooked Venison – an Optical Phenomenon
Journal of Nutritional Health & Food Engineering Research Article Open Access Iridescence in cooked venison – an optical phenomenon Abstract Volume 8 Issue 2 - 2018 Iridescence in single myofibers from roast venison resembled multilayer interference in having multiple spectral peaks that were easily visible under water. The relationship HJ Swatland of iridescence to light scattering in roast venison was explored using the weighted- University of Guelph, Canada ordinate method of colorimetry. In iridescent myofibers, a reflectance ratio (400/700 nm) showing wavelength-dependent light scattering was correlated with HJ Swatland, Designation Professor CIE (Commission International de l’Éclairage) Y%, a measure of overall paleness Correspondence: Emeritus, University of Guelph, 33 Robinson Ave, Guelph, (r=0.48, P< 0.01). Hence, meat iridescence is an optical phenomenon. The underlying Ontario N1H 2Y8, Canada, Tel 519-821-7513, mechanism, subsurface multilayer interference, may be important for meat colorimetry. Email [email protected] venison, iridescence, interference, reflectance, meat color Keywords: Received: August 23, 2017 | Published: March 14, 2018 Introduction balanced pixel hues that have tricked your eyes to appear white; and when we perceive interference colors, complex interference spectra Iridescence is an enigmatic aspect of meat color with some practical trick our eyes again. As the order of interference increases, the colors 1–4 importance for consumers concerned about green colors in meat. appear to change from metallic -

Octopus Consciousness: the Role of Perceptual Richness
Review Octopus Consciousness: The Role of Perceptual Richness Jennifer Mather Department of Psychology, University of Lethbridge, Lethbridge, AB T1K 3M4, Canada; [email protected] Abstract: It is always difficult to even advance possible dimensions of consciousness, but Birch et al., 2020 have suggested four possible dimensions and this review discusses the first, perceptual richness, with relation to octopuses. They advance acuity, bandwidth, and categorization power as possible components. It is first necessary to realize that sensory richness does not automatically lead to perceptual richness and this capacity may not be accessed by consciousness. Octopuses do not discriminate light wavelength frequency (color) but rather its plane of polarization, a dimension that we do not understand. Their eyes are laterally placed on the head, leading to monocular vision and head movements that give a sequential rather than simultaneous view of items, possibly consciously planned. Details of control of the rich sensorimotor system of the arms, with 3/5 of the neurons of the nervous system, may normally not be accessed to the brain and thus to consciousness. The chromatophore-based skin appearance system is likely open loop, and not available to the octopus’ vision. Conversely, in a laboratory situation that is not ecologically valid for the octopus, learning about shapes and extents of visual figures was extensive and flexible, likely consciously planned. Similarly, octopuses’ local place in and navigation around space can be guided by light polarization plane and visual landmark location and is learned and monitored. The complex array of chemical cues delivered by water and on surfaces does not fit neatly into the components above and has barely been tested but might easily be described as perceptually rich. -

INSECTA: LEPIDOPTERA) DE GUATEMALA CON UNA RESEÑA HISTÓRICA Towards a Synthesis of the Papilionoidea (Insecta: Lepidoptera) from Guatemala with a Historical Sketch
ZOOLOGÍA-TAXONOMÍA www.unal.edu.co/icn/publicaciones/caldasia.htm Caldasia 31(2):407-440. 2009 HACIA UNA SÍNTESIS DE LOS PAPILIONOIDEA (INSECTA: LEPIDOPTERA) DE GUATEMALA CON UNA RESEÑA HISTÓRICA Towards a synthesis of the Papilionoidea (Insecta: Lepidoptera) from Guatemala with a historical sketch JOSÉ LUIS SALINAS-GUTIÉRREZ El Colegio de la Frontera Sur (ECOSUR). Unidad Chetumal. Av. Centenario km. 5.5, A. P. 424, C. P. 77900. Chetumal, Quintana Roo, México, México. [email protected] CLAUDIO MÉNDEZ Escuela de Biología, Universidad de San Carlos, Ciudad Universitaria, Campus Central USAC, Zona 12. Guatemala, Guatemala. [email protected] MERCEDES BARRIOS Centro de Estudios Conservacionistas (CECON), Universidad de San Carlos, Avenida La Reforma 0-53, Zona 10, Guatemala, Guatemala. [email protected] CARMEN POZO El Colegio de la Frontera Sur (ECOSUR). Unidad Chetumal. Av. Centenario km. 5.5, A. P. 424, C. P. 77900. Chetumal, Quintana Roo, México, México. [email protected] JORGE LLORENTE-BOUSQUETS Museo de Zoología, Facultad de Ciencias, UNAM. Apartado Postal 70-399, México D.F. 04510; México. [email protected]. Autor responsable. RESUMEN La riqueza biológica de Mesoamérica es enorme. Dentro de esta gran área geográfi ca se encuentran algunos de los ecosistemas más diversos del planeta (selvas tropicales), así como varios de los principales centros de endemismo en el mundo (bosques nublados). Países como Guatemala, en esta gran área biogeográfi ca, tiene grandes zonas de bosque húmedo tropical y bosque mesófi lo, por esta razón es muy importante para analizar la diversidad en la región. Lamentablemente, la fauna de mariposas de Guatemala es poco conocida y por lo tanto, es necesario llevar a cabo un estudio y análisis de la composición y la diversidad de las mariposas (Lepidoptera: Papilionoidea) en Guatemala. -
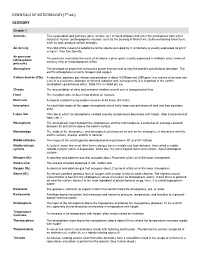
ESSENTIALS of METEOROLOGY (7Th Ed.) GLOSSARY
ESSENTIALS OF METEOROLOGY (7th ed.) GLOSSARY Chapter 1 Aerosols Tiny suspended solid particles (dust, smoke, etc.) or liquid droplets that enter the atmosphere from either natural or human (anthropogenic) sources, such as the burning of fossil fuels. Sulfur-containing fossil fuels, such as coal, produce sulfate aerosols. Air density The ratio of the mass of a substance to the volume occupied by it. Air density is usually expressed as g/cm3 or kg/m3. Also See Density. Air pressure The pressure exerted by the mass of air above a given point, usually expressed in millibars (mb), inches of (atmospheric mercury (Hg) or in hectopascals (hPa). pressure) Atmosphere The envelope of gases that surround a planet and are held to it by the planet's gravitational attraction. The earth's atmosphere is mainly nitrogen and oxygen. Carbon dioxide (CO2) A colorless, odorless gas whose concentration is about 0.039 percent (390 ppm) in a volume of air near sea level. It is a selective absorber of infrared radiation and, consequently, it is important in the earth's atmospheric greenhouse effect. Solid CO2 is called dry ice. Climate The accumulation of daily and seasonal weather events over a long period of time. Front The transition zone between two distinct air masses. Hurricane A tropical cyclone having winds in excess of 64 knots (74 mi/hr). Ionosphere An electrified region of the upper atmosphere where fairly large concentrations of ions and free electrons exist. Lapse rate The rate at which an atmospheric variable (usually temperature) decreases with height. (See Environmental lapse rate.) Mesosphere The atmospheric layer between the stratosphere and the thermosphere. -

Butterfly-Inspired Photonics Reverse Diffraction Color Sequence
COMMENTARY Butterfly-inspired photonics reverse diffraction color sequence Michael H. Bartl1 and plants (3, 14). Structural colors are the Department of Chemistry, and Materials Research Science and Engineering Center, University result of diffraction and specular reflection of Utah, Salt Lake City, UT 84112 of light, in contrast to typical pigments that produce color by light absorption and dif- Biological systems have been an infinite 12). In contrast, in bioinspiration and bio- fuse reflection. Fossil finds date the first reservoir of inspiration ever since humans mimicry, a biological function or activity— examples of structural colors to some 500 million years ago, and the earliest examples started to develop tools and machinery. Just rather than the organism itself—is converted of photonic structures in these fossil records as early scientists and engineers attempted to into an artificial, human-made material or were discovered within insect hairs and spines mimic birds and fish in the development of device. In PNAS, England et al. report an in the form of multilayers and gratings flying machines and submarines, today, new optical micrograting array inspired by a pho- (15). Although these early structures most technologies find their inspiration from bi- tonic structure found in iridescently colored likely were the result of random mutations, ology, such as gecko feet (1), antireflective eye butterfly wings (13). The authors demon- the accompanying coloration effects must lenses (2), iridescent insects (3), and water- strate a micrograting array that not only have presented evolutionary advantages in repellant surfaces (4). Incorporating biologi- mimics the unique diffraction properties of camouflaging, signaling, communicating, cal systems and concepts into technological the biological structure with reversed color- and mimicking (16). -

Ecology, Population Biology and Mortality of Euptoieta Hegesia Cramer (Nymphalidae) on Jamaica
}<mrnal <1 the Lepidopterists' Society 52(1), 1998, 9-39 ECOLOGY, POPULATION BIOLOGY AND MORTALITY OF EUPTOIETA HEGESIA CRAMER (NYMPHALIDAE) ON JAMAICA PHILLIP J. SCHAPPERT] AND JOEL S. SHORE Department of Biology, York University, 4700 Keele Street, North York, Ontario M3J IP3, Canada ABSTRACT. We examine the ecology, population biology and potential sources of mortality of Euptoieta hegesia, a tropical lowland butterfly from Jamaica, using a combina tion of captive rearing, studies of natural populations, and experimental approaches. We provide detailed observations of the life cycle and methods for captive rearing of this spe cies. We assess the relative performance of larvae on primary and secondary hostplants, distribution of larvae on the primary hostplant, hostplant population utilization, and the distribution of E. hegesia on the island. A mark-release-recapture study was conducted to estimate population parameters and we recorded sex, size, age (as estimated by wing wear), and wing damage sustained by the butterflies prior to their initial capture. We pro vide evidence that Tumera ulmifdia is the plimary hostplant of K hegesia on Jamaica and that butterfly population size is not limited by the availability of hostplants. These short lived butterflies appear to be residents of discrete hostplant populations and experience high mortality levels. Females are damaged more frequently, show more total damage and more frequent symmetrical hindwing damage (attributablc to ground-based predators) than do males. Wc compare the results of the population study with available studies of other tropical butterflies and suggest that lowland butterfly population structure and dy namics are Significantly different from that of rainforest species. -

Iridescent Color: from Nature to the Painter's Palette
Downloaded from http://www.mitpressjournals.org/doi/pdf/10.1162/LEON_a_00114 by guest on 25 September 2021 Technical ar T i c l e ce, Tech N e Iridescent Color: From Nature I , Sc to the Painter’s Palette T a b s T r a c T o: Ar nan The shifting rainbow hues of Franziska Schenk iridescence have, until recently, remained exclusive to nature. and Andrew Parker Now, the latest advances in nanotechnology enable the introduction of novel, bio- inspired color-shifting flakes into painting—thereby affording artists potential access to the full spectacle of iridescence. Unfortunately, existing rules rtists have never captured a color as dazzling selves of two distinct types: those of easel painting do not apply A that are stable attributes of material to the new medium; but, as and dynamic as the metallic blue of the Morpho butterfly, which nature inspired the technol- is visible for up to a quarter of a mile. Now, with rapid ad- substances, and those that are “acci- dental,” such as the evanescent col- ogy, an exploration of natural vances in nanoscience and technology, we are beginning to ors of the rainbow and the colors of phenomena can best inform unravel nature’s ingenious manipulation of the flow of light. some birds’ feathers, which change how to overcome this hurdle. Scientific research into natural nanoscale architectures, ca- according to the viewpoint of the Thus, by adopting a biomimetic pable of producing eye-catching optical effects, has led to the spectator [2]. approach, this paper outlines the optical principles underly- development of an ever-expanding range of comparable syn- ing iridescence and provides thetic structures. -

Biological Growth and Synthetic Fabrication of Structurally Colored Materials
Biological growth and synthetic fabrication of structurally colored materials The MIT Faculty has made this article openly available. Please share how this access benefits you. Your story matters. Citation McDougal, Anthony et al. "Biological growth and synthetic fabrication of structurally colored materials." Journal of Optics 21, 7 (June 2019): 073001 © 2019 IOP Publishing Ltd As Published http://dx.doi.org/10.1088/2040-8986/aaff39 Publisher IOP Publishing Version Final published version Citable link https://hdl.handle.net/1721.1/126616 Terms of Use Creative Commons Attribution 3.0 unported license Detailed Terms https://creativecommons.org/licenses/by/3.0/ Journal of Optics TOPICAL REVIEW • OPEN ACCESS Recent citations Biological growth and synthetic fabrication of - Stability and Selective Vapor Sensing of Structurally Colored Lepidopteran Wings structurally colored materials Under Humid Conditions Gábor Piszter et al To cite this article: Anthony McDougal et al 2019 J. Opt. 21 073001 - Iridescence and thermal properties of Urosaurus ornatus lizard skin described by a model of coupled photonic structures José G Murillo et al - Biological Material Interfaces as Inspiration View the article online for updates and enhancements. for Mechanical and Optical Material Designs Jing Ren et al This content was downloaded from IP address 137.83.219.59 on 29/07/2020 at 14:27 Journal of Optics J. Opt. 21 (2019) 073001 (51pp) https://doi.org/10.1088/2040-8986/aaff39 Topical Review Biological growth and synthetic fabrication of structurally colored materials Anthony McDougal , Benjamin Miller, Meera Singh and Mathias Kolle Department of Mechanical Engineering, Massachusetts Institute of Technology, 77 Massachusetts Avenue, Cambridge, MA 02139, United States of America E-mail: [email protected] Received 9 January 2018, revised 29 May 2018 Accepted for publication 16 January 2019 Published 11 June 2019 Abstract Nature’s light manipulation strategies—in particular those at the origin of bright iridescent colors —have fascinated humans for centuries. -

Comparison of Rainforest Butterfly Assemblages Across Three Biogeographical Regions Using Standardized Protocols
The Journal Volume 44: 17-28 of Research on the Lepidoptera ISSN 0022-4324 (PR in T ) THE LEPIDOPTERA RESEARCH FOUNDATION, 4 MA Y 2011 ISSN 2156-5457 (O N L in E ) Comparison of rainforest butterfly assemblages across three biogeographical regions using standardized protocols YVE S Bass ET 1,*, RO D Eas TWOO D 2, LEG I Sam 3, DA V id J. LO hman 2,4, VO J TE ch NOVOT N Y 5, Tim TRE U ER 2, SC OTT E. MI LLER 6, GEORGE D. WE ib LE N 7, NA O mi E. PI ER C E 2, SA R A Y udh Bun Y A VE jch EW in 8, WA T ana SA K ch OOWO N G 8, PI TOO N KO N G N OO 9 and MI G U EL A. OS OR I O -ARE nas 1 1Smithsonian Tropical Research Institute, Apartado 0843-03092, Balboa, Ancon, Panama City, Republic of Panama 2Museum of Comparative Zoology, Harvard University, 26 Oxford Street, Cambridge, MA 02138, USA 3The New Guinea Binatang Research Center, PO Box 604, Madang, Papua New Guinea 4Department of Biology, The City College of New York, The City University of New York, Convent Avenue at 138th Street, New York, NY 10031, USA 5Biology Center of the Czech Academy of Sciences and School of Biological Sciences, University of South Bohemia, Branisovska 31, 370 05 Ceske Budejovice, Czech Republic 6National Museum of Natural History, Smithsonian Institution, Washington, DC 20560-0105, USA 7Bell Museum of Natural History, University of Minnesota, 250 Biological Sciences Center, 1445 Gortner Avenue Saint Paul, Minnesota 55108, USA 8Thai National Parks Wildlife and Plant Conservation Department, 61 Phaholyothin Road, Chatuchak, Bangkok 10900, Thailand 9Center for Tropical Forest Science, Khao Chong Botanical Garden, Tambon Chong, Nayong District, Trang 92170, Thailand [email protected] Abstract. -

Bioinspired Micrograting Arrays Mimicking the Reverse Color Diffraction Elements Evolved by the Butterfly Pierella Luna
Bioinspired micrograting arrays mimicking the reverse color diffraction elements evolved by the butterfly Pierella luna Grant Englanda, Mathias Kollea,b,1, Philseok Kimc, Mughees Khanc, Philip Muñoza, Eric Mazura, and Joanna Aizenberga,c,1 aSchool of Engineering and Applied Sciences, Harvard University, Cambridge, MA 02138; bDepartment of Mechanical Engineering, Massachusetts Institute of Technology, Cambridge, MA, 02139; and cWyss Institute for Biologically Inspired Engineering, Harvard University, Cambridge, MA 02138 Edited by Galen D. Stucky, University of California, Santa Barbara, CA, and approved September 12, 2014 (received for review June 30, 2014) Recently, diffraction elements that reverse the color sequence effect results from the local morphology of individual scales normally observed in planar diffraction gratings have been found within the colored spot on the fore wings (12). The top parts of in the wing scales of the butterfly Pierella luna. Here, we describe the scales are curled upward, orienting lines of periodically the creation of an artificial photonic material mimicking this re- arranged cross-ribs perpendicular to the wing surface. (Fig. 1 C verse color-order diffraction effect. The bioinspired system con- and D). Light incident at an angle onto the curled parts of the sists of ordered arrays of vertically oriented microdiffraction scales is diffracted by the cross-rib structure acting as a diffrac- gratings. We present a detailed analysis and modeling of the cou- tion grating, with a periodicity of ∼400 nm. The alignment of the pling of diffraction resulting from individual structural compo- grating perpendicular to the surface results in the reverse color nents and demonstrate its strong dependence on the orientation sequence that can be observed in angularly resolved reflection of the individual miniature gratings. -

Journal of Avian Biology JAV-02474 Mills, L
Journal of Avian Biology JAV-02474 Mills, L. J., Wilson, J. D., Lange, A., Moore, K., Henwood, B., Knipe, H., Chaput, D. L. and Tyler, C. R. 2020. Using molecular and crowd-sourcing methods to assess breeding ground diet of a migratory brood parasite of conservation concern. – J. Avian Biol. 2020: e02474 Supplementary material Supplementary materials DNA extraction Ethanol was removed from the samples by freeze-drying to prevent loss of DNA from the vials, and dried samples were stored at -80 °C until DNA extraction. DNA was extracted using a precipitation and re-suspension method adapted from Bramwell et al. (1995), with modifications recommended by Lever et al. (2015). Lysis buffer (30 mM Tris, 30 mM EDTA, pH 8) was added at room temperature and mixed by vortex. Samples were Quickly re-frozen in liQuid nitrogen to lyse cells and then briefly thawed in a 37 °C water bath. A mixture of two different beads (0.2 g ceramic beads 1.4 mm and 0.3 g garnet beads 0.7 mm) was added and samples were shaken at 30 Hz for 3 x 40 s using a TissueLyser II (Qiagen, Hilden Germany). To 19-parts sample solution, 1 part SDS solution (10% w/v) and 0.1 parts proteinase K (20 mg/ml) were added, and samples incubated for 4 h in a shaking incubator at 55 °C to continue lysis and protein digestion. Sample temperature was raised to 65 °C and to each 5 part sample solution, 1 part of 5 M NaCl solution was added and mixed by inversion, then 0.8 parts warm CTAB solution (hexadecyltrimethylammonium bromide, 10% w/v) added and mixed.