New Host Records for Euploea Core Corinna (Macleay)
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Species Diversity and Community Structure of Butterfly in Urban Forest Fragments at Lucknow, India
Journal of Applied and Natural Science 10 (4): 1276-1280 (2018) ISSN : 0974-9411 (Print), 2231-5209 (Online) journals.ansfoundation.org Species diversity and community structure of butterfly in urban forest fragments at Lucknow, India Ashok Kumar* Article Info Department of Zoology, BSNVPG College (Lucknow University), Lucknow (U.P.), India DOI:10.31018/jans.v10i4.1908 Satyapal Singh Rana Received: September 26, 2018 Department of Zoology, S. M. P. Govt. Girls P.G. College, Meerut (U.P.), India Revised: November 18, 2018 Accepted: November 27, 2018 *Corresponding author. E-mail: ashokbsnv11gmail.com Abstract The survey was carried out between September 2015-August 2016 in five different locali- How to Cite ties in Lucknow like Bijli Pasi Quila, Smriti Upvan, Vanasthali Park, Butchery Ground and Kumar, A. and Rana, S.S. BSNVPG College Campus, Lucknow, 26.84’N latitude and 80.92’E longitude, is located at (2018). Species diversity an elevation of 126 meters above sea level and in the plain of northern India. Its location and community structure of is responsible for the diverse weather patterns and climate change. The butterfly in urban forest region has tropical dry equable climate having three main seasons; cold, hot and rainy fragments at Lucknow, season. Temperature of the city ranges from 23.8- 45.8°C in summer and 4.6-29.7°C in India. Journal of Applied winter. During the study, butterflies were collected mainly with the help of circular aerial and Natural Science, 10 net, which were then placed in killing jar. Killed butterflies were stored in the insect box by (4): 1276-1280 proper pinning them for identification. -
Appendix 1 Table A1
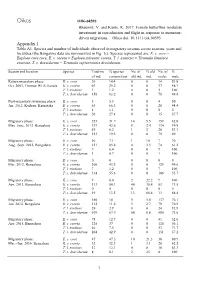
Oikos OIK-04593 Bhaumik, V. and Kunte, K. 2017. Female butterflies modulate investment in reproduction and flight in response to monsoon- driven migrations. – Oikos doi: 10.1111/oik.04593 Appendix 1 Table A1. Species and number of individuals observed in migratory swarms across seasons, years and localities (the Bangalore data are summarized in Fig. 1c). Species represented are: E. c. core = Euploea core core, E. s. coreta = Euploea sylvester coreta, T. l. exoticus = Tirumala limniace exoticus, T. s. dravidarum = Tirumala septentrionis dravidarum. Season and location Species Total no. % species No. of % old No. of % of ind. composition old ind. ind. males male Return migratory phase: E. c. core 26 10.4 0 0 14 53.8 Oct. 2001, Chinnar WLS, Kerala E. s. coreta 63 25.2 0 0 37 58.7 T. l. exoticus 3 1.2 0 0 3 100 T. s. dravidarum 158 63.2 0 0 78 49.4 Post-migratory swarming phase: E. c. core 5 5.3 0 0 4 80 Jan. 2012, Kodagu, Karnataka E. s. coreta 63 66.3 0 0 28 44.4 T. l. exoticus 1 1.1 0 0 0 0 T. s. dravidarum 26 27.4 0 0 15 57.7 Migratory phase: E. c. core 253 31.9 14 5.5 159 62.8 May–June, 2012, Bengaluru E. s. coreta 337 42.6 8 2.4 134 39.8 T. l. exoticus 49 6.2 1 2 26 53.1 T. s. dravidarum 153 19.3 0 0 75 49 Migratory phase: E. c. core 10 7.1 1 10 7 70 Aug.–Sept. -

Euploea Species Are Unpalatable to Their Otherwise Enemies
An introduction to Crows of the World, the Euploeas (Lepidoptera : Danainae) – Peter Hendry The Euploea belong to the subfamily Danainae of the butterfly family Nymphalidae, although some authors place them in a family of their own, Danaidae. They are known as Crows, because of their mostly dark colouration, generally dark brown with some white markings. Some Crows have a blue or purplish sheen over part of their wings. With one endemic species on the Seychelles and another on the Mascarene Islands, in the Indian Ocean, the remainder of the genus ranges from India and South-east Asia (from Sikkim, Tibet and Afghanistan in the east) as far north as the Ryukyu Islands of southern Japan, throughout the Philippines and the Indonesian Archipelago, to New Guinea, the Bismarcks, Solomons, northern and eastern Australia, and the Islands associated with New Caledonia, the New Hebrides (Vanuatu), Fiji, Samoa, Tonga and as far eastwards as Niue, the Cook Islands and the Society Islands. (Parsons Fig. 1 1999) Fig.1. The larvae feed on plants in the Apocynaceae, Asclepiadaceae and Moraceae plant families, many of which are poisonous to most animals. Most of these plants extrude milky latex when cut. Caterpillars of the Common Crow, Euploea core corinna (Fig. 16) sever leaf veins prior to feeding on their latex-bearing host plants, which restricts the flow of latex at feeding sites (Clarke and Zalucki, 2001). This has been observed for other species and genera. Over 100 species of Lepidoptera, Coleoptera and Orthoptera cut veins on host plants (Helmus and Dussourd, 2003). By feeding on toxic plants the larvae, pupae and adults of some Euploea species are unpalatable to their otherwise enemies. -

Observations on the Ecology of Euploea Core Corinna (Nymphalidae) with Special Reference to an Overwintering Population
Journal of the Lepidopterists' Society 35(2), 1981, 106-119 OBSERVATIONS ON THE ECOLOGY OF EUPLOEA CORE CORINNA (NYMPHALIDAE) WITH SPECIAL REFERENCE TO AN OVERWINTERING POPULATION R. L. KITCHING AND M. P. ZALUCKI School of Australian Environmental Studies, Griffith University, Nathan, QLD 4111, Australia ABSTRACT. A study of a sub-tropical overwintering aggregation of the common crow butterfly, Euploea core corinna (Nymphalidae: Danainae), has been made on the campus of Griffith University, Brisbane, Australia. Observations on the temporal and spatial phenology of the aggregation, together with the sex ratio, reproductive status, individual fat content and physical condition of butterflies within the aggregation are presented. The aggregation was restricted to the upper reaches of a gully on the campus between mid-May and late July. Physical conditions in the gully (temperature, humid ity and wind) were less severe than in adjacent areas. The butterflies in the gully were reproductively inactive. We suggest that such aggregations, apart from avoiding severe winter conditions, may also serve as mating concourses at the end of the winter season. The common crow butterfly, Euploea core (Cramer), is widely dis tributed in the Indo-Australian region and is represented in Australia by the subspecies, corinna (W. S. Macleay) (Fig. 1). Recent general ac counts of its life history are given by McCubbin (1971) and Common & Waterhouse (1972). These authors summarize information on food plants, phenology, behavior and morphology, principally collated from the observations of field naturalists. McCubbin (1971) alluded to the formation of overwintering aggregations of this species in shel tered coastal sites and offshore islands in the tropics and subtropics of northern and eastern Australia. -

Studies on Butterfly Diversity in Adichanalloor Village, Kollam
Journal of Entomology and Zoology Studies 2017; 5(5): 73-81 E-ISSN: 2320-7078 P-ISSN: 2349-6800 JEZS 2017; 5(5): 73-81 Studies on butterfly diversity in Adichanalloor © 2017 JEZS Village, Kollam District, Kerala Received: 11-07-2017 Accepted: 12-08-2017 Lekshmi Priya Lekshmi Priya, Varunprasath Krishnaraj, Janaranjini, Sutharsan and Department of Zoology, PSG Lakeshmanaswamy College of Arts and Science, Coimbatore, Tamil Nadu, India Abstract Varunprasath Krishnaraj The present investigation was carried out to study butterfly diversity in Adichanalloor Village, Kollam Department of Zoology, PSG district in Kerala, for the period of November 2016 to March 2017. Results showed that 79 species of College of Arts and Science, butterflies representing 5 major families were recorded. Family Nymphalidae showed the maximum Coimbatore, Tamil Nadu, India number of species followed by Lycanidae 13 species, Papilionidae 10 species, Pieridae 9 species and Hesperiidae 7 species. Among these families abundance of butterfly species in maximum in garden area Janaranjini (GI) with 21 species, followed by agrifield (GIII) (17 species), pond region (GV) (16 species), grassland Department of Zoology, PSG College of Arts and Science, (GII) (13 species) and shrubs and herbs (GIV) (12 species).Based on IUCN list, 49 species were Coimbatore, Tamil Nadu, India common(C), 27 species, uncommon (UC) and 3 species under rare category. According to monthly wise distribution of butterflies, maximum numbers of butterflies were recorded in November (32 species) Sutharsan followed by a December (21 species), January (12 species) and least in the month of March (8 species). Department of Zoology, PSG College of Arts and Science, Keywords: distribution, butterflies, Adichanalloor village, Kollam district, abundance. -

Sharma Ethno Ferns
CASE REPORT ZOOS' PRINT JOURNAL 18(2): 1003-1006 BUTTERFLIES OF SIRUVANI FORESTS OF WESTERN GHATS, WITH NOTES ON THEIR SEASONALITY P.R. Arun Salim Ali Centre for Ornithology and Natural History, Moongilpallam, Anaikatty, Coimbatore, Tamil Nadu 641108, India E-mail: [email protected] Abstract Study area The seasonal pattern in the abundance of butterflies The moist deciduous forests of Siruvani falling between 76o41'- belonging to the families, Papilionidae, Pieridae and 45' E and 10o56'-58' N in the Boluvampatti Reserve Forest located Nymphalidae was monitored during the period of in the foothills of Western Ghats about 35km west of Coimbatore, September 1994 to August 1996 in the moist deciduous Tamil Nadu, southern India was the selected study area. The forest of Siruvani, using the transect counting method. area receives good rainfall during both the north-east and south- Seventy-five species of butterflies belonging to 49 genera west monsoons. The mean annual rainfall of the area during the were recorded from the area during the study. Of these study period was 2092mm, much higher than that generally 53 were encountered along the transect. The present received by moist deciduous zones (Champion & Seth, 1968), paper also provides an account of their seasonality owing to the closeness of the study area to one of the core zone pattern. evergreen forests of the Nilgiri Biosphere Reserve. Keywords The transect selected for the study was about four kilometre off Butterflies, Lepidoptera, seasonal pattern, Siruvani, the forest check post at Sadivayal on the road towards the Water Nilgiri Biosphere Reserve, Western Ghats, southern Filtering plant of TWAD (Tamilnadu Water Supply and Drainage) India Board. -

Download Download
PLATINUM The Journal of Threatened Taxa (JoTT) is dedicated to building evidence for conservaton globally by publishing peer-reviewed artcles online OPEN ACCESS every month at a reasonably rapid rate at www.threatenedtaxa.org. All artcles published in JoTT are registered under Creatve Commons Atributon 4.0 Internatonal License unless otherwise mentoned. JoTT allows allows unrestricted use, reproducton, and distributon of artcles in any medium by providing adequate credit to the author(s) and the source of publicaton. Journal of Threatened Taxa Building evidence for conservaton globally www.threatenedtaxa.org ISSN 0974-7907 (Online) | ISSN 0974-7893 (Print) Communication Butterflies of the myristica swamp forests of Shendurney Wildlife Sanctuary in the southern Western Ghats, Kerala, India Prabhakaran Chandrika Sujitha, Gopal Prasad & Kalesh Sadasivan 26 February 2019 | Vol. 11 | No. 3 | Pages: 13320–13333 DOI: 10.11609/jot.4399.11.3.13320-13333 For Focus, Scope, Aims, Policies, and Guidelines visit htps://threatenedtaxa.org/index.php/JoTT/about/editorialPolicies#custom-0 For Artcle Submission Guidelines, visit htps://threatenedtaxa.org/index.php/JoTT/about/submissions#onlineSubmissions For Policies against Scientfc Misconduct, visit htps://threatenedtaxa.org/index.php/JoTT/about/editorialPolicies#custom-2 For reprints, contact <[email protected]> The opinions expressed by the authors do not refect the views of the Journal of Threatened Taxa, Wildlife Informaton Liaison Development Society, Zoo Outreach Organizaton, or any of the -

Butterflies in Wetlands of Kakadu National Park, Northern Australia
BUTTERFLIES IN WETLANDS OF KAKADU NATIONAL PARK, NORTHERN AUSTRALIA Butterflies can be found invertebrates play in helping us to identify and in various habitats preserve critical habitats under threat. Butterflies can throughout Kakadu be used as indicators for monitoring climate change National Park, from or pollution, or identified as rare or threatened KAKADU BUTTERFLIES shady, tranquil vine species themselves needing conservation. We thickets to the diverse anticipate that further analysis of invertebrate savanna woodland and biodiversity in Kakadu’s wetland habitats will vast, spectacular illustrate the importance of these animals and their wetlands. At certain potential as indicators of environmental change. times of the year some However, first we need further information on the species congregate in ecology and habitat preferences of such species. This great numbers in cool, analysis of butterflies is an example of this effort – an Photo by Caroline Camilleri monsoon rainforest effort that extends across the landscape and habitats pockets. Others frequent the shady creeks and springs of the Kakadu region. The Environmental Research This publication is dedicated to Murrumbur that create deep gorges in the escarpment and flow Institute of the Supervising Scientist is contributing clan member Mick Alderson. Mick inspired us out into the savanna woodland. Some butterflies to this effort in collaboration with local people and prefer to inhabit paperbark forests, mangrove swamps land managers. to produce this guide by suggesting a pamphlet and estuaries along the coast. The preference of butterflies for particular habitats about butterflies. Although he will never see There are about 100 species of butterflies is often closely linked to the adult or larval food this finished product, we trust that it will be inhabiting Kakadu. -

RECORD of NATURAL ENEMIES of FEW BUTTERFLY SPECIES AMIDST AGRICULTURE ECOSYSTEMS of CHAMARAJANAGAR DISTRICT, KARNATAKA, INDIA Santhosh S., S
Santhosh & Basavarajappa RJLBPCS 2017 www.rjlbpcs.com Life Science Informatics Publications Original Research Article DOI - 10.26479/2017.0205.02 RECORD OF NATURAL ENEMIES OF FEW BUTTERFLY SPECIES AMIDST AGRICULTURE ECOSYSTEMS OF CHAMARAJANAGAR DISTRICT, KARNATAKA, INDIA Santhosh S., S. Basavarajappa* Entomology Laboratory, DOS in Zoology University of Mysore, Manasagangotri, Mysore-570 006, India ABSTRACT: Field studies were carried out by following various standard methods to record the natural enemies of 19 butterfly species during Rabi and Kharif crops amidst agriculture ecosystems of Chamarajanagar District (110401 to 120481 NL and 740521 to 760071 EL) in south Deccan plateau at southern tip of Karnataka State, India. The oviposition and pupation of butterfly species were critically observed for the parasites and predators attack. The death of egg, pupa, larva and adult butterfly species due to parasitoids and spiders was recorded. The parasitoid, Brachymeria jambolana (Hymenoptera: Chalcididae) was more troublesome to the pupa of G. doson and E. core. Around 25 to 27 B. jambolana adult individuals were collected respectively from the pupa of G. doson and E. core. Moreover spider species viz., Selenopes, Zygeilla, Argiope, Hippasa, and Rhene become common predators to 16 butterfly species in this part of the State. Thus, parasitoids and spiders infestation could cause loss to several insect species including protected species like E. core. As natural enemy’s interferences is natural in the wild/open cultivated conditions, it could become one of the major reasons to reduce local butterfly diversity. Hence, there is a dire need to understand about the interaction of parasitoid species with butterfly species which are under threat and whose presence is very essential for the restoration of local biodiversity. -

Biodiversity of One of the Mishing Villages of Assam with an Emphasis of the Traditional Knowledge and Use of the Wild Plants
International Journal for Environmental Rehabilitation and Conservation ISSN: 0975 — 6272 IX (2): 80— 86 www.essence-journal.com Original Research Article Biodiversity of one of the Mishing villages of Assam with an emphasis of the traditional knowledge and use of the wild plants Dutta Mary1, Basumatari Mallika1, Borah Darshana1, Gogoi Lina2 1Balipara Tract and Frontier Foundation, Sonitpur, Assam, India 2Department of Energy, Tezpur University, Napaam, Tezpur, Assam, India Corresponding Author: [email protected] A R T I C L E I N F O Received: 09 June 2018 | Accepted: 04 October 2018 | Published Online: 31 December 2018 DOI: 10.31786/09756272.18.9.2.212 EOI: 10.11208/essence.18.9.2.212 Article is an Open Access Publication. This work is licensed under Attribution-Non Commercial 4.0 International (https://creativecommons.org/licenses/by/4.0/) ©The Authors (2018). Publishing Rights @ MANU—ICMANU & ESSENCE—IJERC. A B S T R A C T The Mishing tribe is one of the hordes of the Mongoloid groups of people occupying the hills and valleys of northeastern India. Mishing or Miri tribe inhabiting the districts of Dhemaji, North Lakhimpur, Sonitpur, Tinsukia, Dibrugarh, Sibsagar, Jorhat and Golaghat of Assam, Northeast India. Many of the wild plants are used in the livelihood strategies of the tribal people. The present study highlights the biodiversity of one of the Mishing tribe villages of Assam with an emphasis on the traditional knowledge and use of these wild plants. A total of 31 plant and 19 bird species were recorded in the present study. In our survey, 67 species of butterflies of various families are reported in the village. -

Butterfly Diversity in Lateritic Biotope of Kavvayi River Basin, Kerala, India
Current World Environment Vol. 12(1), 132-141 (2017) Butterfly Diversity in Lateritic Biotope of Kavvayi River Basin, Kerala, India DHANEESH BHASKAR, M. P. PREJITH, K. P. RAJKUMAR, C. J. ALEX1, T. S. PRASAD and K. A. SREEJITH* Forest Ecology and Biodiversity Conservation Division, Kerala Forest Research Institute, Peechi, Thrissur, Kerala. Corresponding author Email: [email protected] http://dx.doi.org/10.12944/CWE.12.1.16 (Received: March 04, 2017; Accepted: April 11, 2017) ABSTRACT A study on butterfly diversity of lateritic biotopes of Kavvayi River Basin was conducted during February 2013 to January 2015. The study area represents diverse habitats that include sacred groves, laterite hills, riparian ecosystem, and kanams. A total of 140 species butterflies were recorded from the study area, among which the highest number of butterfly species were from the family Nymphalidae (48 species), followed by Hesperiidae (32 species), Lycaenidae (27 species), Papilionidae and Pieridae with 16 species each and 1 from the family Riodinidae. The present study revealed the faunal richness of the unique ecosystems and microhabitats in lateritic biotopes in terms of butterfly diversity. The study also highlights conservation significance of the area which is under severe human pressure including mining, habitat fragmentation and change in the land-use system. Keywords: Laterite ecosystem, Host plant, Butterfly diversity, Northern Kerala. INTRODUCTION reported butterfly species of Western Ghats, 316 species of butterflies were recorded from Kerala10. As The midland exposed laterite biotope per the review of literature, a large number of studies associated with diverse ecosystems and on diversity and distribution of butterflies were done microhabitats represents the most imposing and in the protected areas of Kerala includes; Mathew & extremely threatened topographical floristic and Rahmathulla11 who reported 100 species from Silent faunistic features in the northern part of Kerala. -

Biodiversity Mapping Survey/Study in Kaziranga National Park (KNP)
Biodiversity Mapping Survey/Study in Kaziranga National Park (KNP) Insects Chapter Final Report Prepared by Mr. Rahul Khot Entomologist 1 1. Introduction In terrestrial ecosystems, insects play a vital function as herbivores, pollinators, predators and parasites (Seimann and Weisser 2004). Insects are considered to pollinate nearly 70% of crop plants worldwide and over 98% of trees (Klein et al. 2006). Insects inhabit every terrestrial habitat on the planet and play a major role in the evolution and maintenance of biotic communities. They are the primary pollinators of flowering plants; they are important consumers and recyclers of decaying organic matter; and they are integral components in the food-webs of vertebrates and other invertebrates. For these reasons, and many others, the study of insects and their relatives is of increasing importance as society faces increased challenges to preserve and enhance environmental quality, reduce pesticide usage, increase crop productivity, control food costs, and increase trade in the global community. The damage cause by pest species is far outweighed by the positive effects of beneficial species. Pollinators ensure the production of fruit, parasitoids and predators help control pest species, some species contain chemicals of pharmaceutical value, and a large number of species contribute to the decomposition and recycling of dead and decaying matter. The Biodiversity Strategy and Action Plan for Northeast Ecoregion states that 3,624 species of insects are recorded from the region (Tripathi and Barik 2003). Butterflies and moths are by far the best-studied invertebrate organisms in Northeast India, and the region contributes the maximum number of species for the group in the country.