A New Bathyal Shark Fauna from the Pleistocene Sediments of Fiumefreddo (Sicily, Italy)
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Squaliform Shark Teeth of the Genus Centroselachus from the Miocene of Japan
Jour. Geol. Soc. Japan, Vol. 114, No. 10, p. 536-539, October 2008 Squaliform shark teeth of the genus Centroselachus from the Miocene of Japan Hideshi Suzuki *† Received April 21, 2008. Accepted September 24, 2008. * Graduate school of Natural Science and Technology, Division of Environmental Science and Engineering, Kanazawa Uni- versity, Kakuma, Kanazawa 920-1192, Japan † Tateshina Senior High School, 3652, Ashida, Tateshina Town, Kitasaku Gun, Nagano 384-2305 Japan. Abstract: Newly found fossil shark teeth of a sleeper shark are described. This is probably an undescribed species of the genus Centroselachus belonging to the Family Somniosidae. These fossil teeth were discovered from the Middle Miocene Iseyama Formation(Northern Fossa Magna Region), Ueda City, Nagano Prefecture, central Japan. These teeth indicate the shape of a part of a symphysial tooth row, which belong to a left side of Fig. 1. Map showing the fossil locality of Centroselachus sp. ※: parasymphysial teeth and a left first anterior tooth. 138 °18 ′00.11 ″E, 36 °25 ′12.05 ″N, using the topographical map Judging from the main characters of a parasymphysial of“Sanada”, scale 1: 25,000, published by Geographical Survey tooth, such as distal and mesial blades presented both Institute. joining in a notch, it is considered that the tooth differs from teeth of the genus Centroscymnus. This fossil is identified as the genus Centroselachus sp.. This paper constitutes the first fossil record of the genus Cen- troselachus from the Miocene of Japan. Key words: Centroselachus, Somniosidae, Middle Miocene, Iseyama Formation, Northern Fossa Magna Region, Nagano Prefecture, symphysial tooth row. Introduction The purpose of this study is to report newly found fossil shark teeth of the sleeper shark of the genus Centroselachus from the Miocene of Japan. -

MD Shark Catch Card
Live Releases Dead Releases TODAY’S DATE: DATE: TODAY’S MM Write the # of sharks released alive in the Live Releases Box and the # released dead Maryland Shark in the Dead Release Box. DD Catch Card YYYY Today’s Date: MM DD YYYY Species (see reverse for list): If you cannot ID, you MUST release it Shark TAG #: Shark TAG HMS PERMIT #: Was this fish caught during a tournament? Yes / No Tournament Name: Fork Length Tagging Location Reporting Station: *Measurement Description Shark Tag #: * Measure in a straight line from the tip of the (Obtain at Reporting Station) Sex (circle one): M / F snout to the fork of the tail. *Fork Length (inches): **Weight (lbs) ** Scale weight ONLY. Do not record estimated weight. Complete this section if fishing from a vessel: NOTE: Most sharks landed in Maryland must have Vessel Name: a LANDING TAG affixed before removal from the vessel. Tags are available at all SHARK REPORTING STATIONS. To obtain a LANDING TAG, captains or Registration # (State ID): operators of the permitted vessel, or licensed angler if fishing from shore, must complete and submit a Permit Holders Name: catch card for every shark landed. For a complete Atlantic HMS list of sharks that you must report, please see the Permit #: reverse side of this card. Trip Type: Charter / Private / Headboat This information collected is approved under OMB (Circle One) Control # 0648-0328 and expires on 07/31/2019 Live Releases Dead Releases TODAY’S DATE: DATE: TODAY’S MM Write the # of sharks released alive in the Live Releases Box and the # released dead Maryland Shark in the Dead Release Box. -

Integrating Multiple Chemical Tracers to Elucidate the Diet and Habitat of Cookiecutter Sharks Aaron B
www.nature.com/scientificreports OPEN Integrating multiple chemical tracers to elucidate the diet and habitat of Cookiecutter Sharks Aaron B. Carlisle1*, Elizabeth Andruszkiewicz Allan2,9, Sora L. Kim3, Lauren Meyer4,5, Jesse Port6, Stephen Scherrer7 & John O’Sullivan8 The Cookiecutter shark (Isistius brasiliensis) is an ectoparasitic, mesopelagic shark that is known for removing plugs of tissue from larger prey, including teleosts, chondrichthyans, cephalopods, and marine mammals. Although this species is widely distributed throughout the world’s tropical and subtropical oceanic waters, like many deep-water species, it remains very poorly understood due to its mesopelagic distribution. We used a suite of biochemical tracers, including stable isotope analysis (SIA), fatty acid analysis (FAA), and environmental DNA (eDNA), to investigate the trophic ecology of this species in the Central Pacifc around Hawaii. We found that large epipelagic prey constituted a relatively minor part of the overall diet. Surprisingly, small micronektonic and forage species (meso- and epipelagic) are the most important prey group for Cookiecutter sharks across the studied size range (17–43 cm total length), with larger mesopelagic species or species that exhibit diel vertical migration also being important prey. These results were consistent across all the tracer techniques employed. Our results indicate that Cookiecutter sharks play a unique role in pelagic food webs, feeding on prey ranging from the largest apex predators to small, low trophic level species, in particular those that overlap with the depth distribution of the sharks throughout the diel cycle. We also found evidence of a potential shift in diet and/or habitat with size and season. -

Working Document on Elasmobranch Fishes 2001-2011 Final
Working Document presented to the Working Group on Elasmobranch Fishes ICES WGEF, - Lisbon 18-26 June 2012 ——————————————————————————————————— Results on main elasmobranch species captured during the 2001-2011 Porcupine Bank (NE Atlantic) bottom trawl surveys S. Ruiz-Pico (1), F. Baldó (2), F. Velasco (1), O. Fernández-Zapico (1), C. Rodríguez-Cabello (1) (1) Instituto Español de Oceanografía, Centro Oceanográfico de Santander P.O. Box 240, 39080 Santander, Spain (2) Instituto Español de Oceanografía, Centro Oceanográfico de Cádiz Puerto pesquero, Muelle de Levante s/n, 11006 Cádiz, Spain Abstract This working document presents the results on nine of the most important elasmobranch species of the Porcupine bank Spanish surveys during the last decade (2001-2011). The shark species more abundant in these surveys in biomass terms were blackmouth catshark (Galeus melastomus), birdbeak dogfish (Deania calcea), Knifetooth dogfish (Scymnodon ringens), velvet belly lantern shark (Etmopterus spinax), lesser spotted dogfish (Scyliorhinus canicula) and bluntnose sixgill shark (Hexanchus griseus); while sandy ray (Leucoraja circularis), cuckoo ray (Leucoraja naevus) and common skate (Dipturus spp. / Dipturus cf. flossada) were the more frequent Rajidae. Biomass, distribution and length ranges were analysed. Many of these species occupy mainly the deep areas covered in the survey, especially D. calcea and S. ringens. Introduction The Porcupine Bank bottom trawl survey has been carried out annually since 2001 to provide data and information for the assessment of the commercial fish species in the area (ICES divisions VIIc and VIIk) (ICES, 2010, 2011a). The aim of this working document is to update the results (abundance indices, length frequency distributions and geographic distributions) on the most common elasmobranch species in Porcupine bottom trawl surveys following the results presented previously (Velasco et al. -

Pdf 1005.43 K
Egyptian Journal of Aquatic Biology & Fisheries Zoology Department, Faculty of Science, Ain Shams University, Cairo, Egypt. ISSN 1110 – 6131 Vol. 23(4): 503 – 519 (2019) www.ejabf.journals.ekb.eg New records, conservation status and pectoral fin description of eight shark species in the Egyptian Mediterranean waters. Walaa M. Shaban* and Mohamed A. M. El-Tabakh Zoology Department, Marine Biology and Fish Sciences section, Faculty of Science, Al-Azhar University, Cairo, Egypt *Corresponding author: [email protected] ARTICLE INFO ABSTRACT Article History: For a taxonomical purpose, the present study described and analyzed the Received: Oct. 12, 2019 pectoral fin shape and measurements of shark specimens, collected from the Accepted: Nov. 6, 2019 Egyptian Mediterranean Sea waters at Alexandria, during the period from Online: Nov. 11, 2019 May 2017 to June 2018. Morphology and morphometric fin characters were _______________ used taxonomically to differentiate between shark species via photo program analysis. After confirming the identification of sharks, a list of shark species Keywords: in the Egyptian Mediterranean waters was given with emphasize on new Shark record species as well as the conservation status of each species. Pectoral fin Results showed that the collected specimens belong to eight species Morphometric analysis from different six families belonging to four orders. Species-list of sharks in New Record this study including; Heptranchias perlo, Hexanchus griseus, Squalus Conservation status megalops, Centrophorus uyato, Oxynotus centrina, Squatina squatina, Mediterranean Sea Isurus oxyrinchus and Isurus paucus. By comparing the present findings Egypt with the previous studies, three shark species out of these eight species were considered as new records in the Egyptian Mediterranean waters. -

WD Porcupine Elasmobranch Fish 2010
Working Document presented to the Working Group on Elasmobranch Fishes ICES WGEF, - Açores 22-29 June 2010 Results on main elasmobranch species captured during the 2001-2009 Porcupine Bank (NE Atlantic) bottom trawl surveys F. Velasco, M. Blanco and F. Baldó Instituto Español de Oceanografía Centro Oceanográfico de Santander P.O. Box 240 39080 Santander, Spain [email protected] Abstract This paper presents the results on nine of the most important elasmobranch fish species of the nine years (2001-2009) of the Porcupine bank Spanish surveys. The main species in biomass terms in Porcupine bank bottom trawl survey, in decreasing biomass abundance order, are: blackmouth catshark, birdbeak dogfish, velvet belly, Knifetooth dogfish, lesser spotted dogfish, bluntnose sixgill shark, sandy ray, cuckoo ray and common skate. Many of these species occupy mainly the deep areas covered in the survey, especially birdbeak dogfish, knifetooth dogfish and velvet belly. Less confined to deeper grounds are blackmouth catshark and sandy ray, while lesser spotted dogfish and cuckoo ray inhabit mainly the shallower grounds close to the Irish shelf or the central mound in the bank. Length distributions of these species along the survey series are also presented and discussed. The few available data on siki sharks from the Porcupine bank survey are also summarized. 1. Introduction Since 2001 a Spanish bottom trawl survey has been carried out annually in the areas surrounding the Porcupine Bank (ICES Divisions VIIc and VIIk) to study the distribution, relative abundance and biological parameters of commercial fish in the area (ICES, 2010a). The main target species for this survey series are hake, monkfish, white anglerfish and megrim, which abundance indices are estimated by age (Velasco et al ., 2005; Velasco et al ., 2007). -
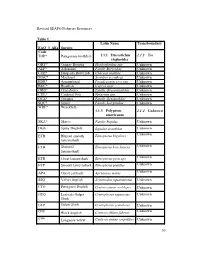
Revised SEAFO Fisheries Resources Table 1. . FAO 3 Alfa Species Code
Revised SEAFO Fisheries Resources Table 1. Latin Name Transboundary FAO 3 Alfa Species Code TOP* Patagonian toothfish 1.1.1 Dissostichus 1.1.2 Yes eleginoides ORY* Orange Roughy Hoplosthethus spp Unknown ALF* Alfonsino Family Berycidae Unknown CGE* Deep-sea Red Crab Chaceon maritae Unknown MAC* Mackerel Scomber scombrus Unknown EDR* Armourhead Pseudopentaceros spp. Unknown BOC* Boarfish Capros aper Unknown ORD* Oreo dories Family Oreosomatidae Unknown CDL* Cardinal Fish Epigonus spp. Unknown OCZ* Octopus Family Octopodidae Unknown SQC* Squid Family Loliginidae Unknown WRF* Wreckfish 1.1.3 Polyprion 1.1.4 Unknown americanus SKA* Skates Family Rajidae Unknown DGS Spiny Dogfish Squalus acanthias Unknown Unknown ETB Blurred smooth Etmopterus bigelowi lanternshark ETH Shorttail Etmopterus brachyurus Unknown lanternshark ETR Great lanternshark Etmopterus princeps Unknown ETP Smooth lanternshark Etmopterus pusillus Unknown Unknown APA Ghost catshark Apristurus manis SSQ Velvet dogfish Scymnodon squamulosus Unknown CYO Portuguese Dogfish Centroscymnus coelolepis Unknown GUQ Leafscale Gulper Centrophorus squamosus Unknown Shark GUP Gulper Shark Centrophorus granulosus Unknown CFB ǂ Black dogfish Centroscyllium fabricii Unknown CYP ǂ Longnose velvet Centroscymnus crepidater Unknown 30 dogfish CYY Unknown ǂ Shortnose velvet Centroscymnus dogfish cryptacanthus SCK ǂ Kitefin shark Dalatias licha Unknown ETE ǂ Etmopterus compagnoi Unknown ETI ǂ Broadbanded Etmopterus gracilispinis Unknown lanternshark ETM ǂ Unknown Southern Etmopterus granulosus lanternshark -

WKSHARK6 Report 2020
WORKSHOP ON THE DISTRIBUTION AND BYCATCH MANAGEMENT OPTIONS OF LISTED DEEP-SEA SHARK SPECIES (WKSHARK6) VOLUME 2 | ISSUE 76 ICES SCIENTIFIC REPORTS RAPPORTS SCIENTIFIQUES DU CIEM ICES INTERNATIONAL COUNCIL FOR THE EXPLORATION OF THE SEA CIEM CONSEIL INTERNATIONAL POUR L’EXPLORATION DE LA MER International Council for the Exploration of the Sea Conseil International pour l’Exploration de la Mer H.C. Andersens Boulevard 44-46 DK-1553 Copenhagen V Denmark Telephone (+45) 33 38 67 00 Telefax (+45) 33 93 42 15 www.ices.dk [email protected] The material in this report may be reused for non-commercial purposes using the recommended cita- tion. ICES may only grant usage rights of information, data, images, graphs, etc. of which it has owner- ship. For other third-party material cited in this report, you must contact the original copyright holder for permission. For citation of datasets or use of data to be included in other databases, please refer to the latest ICES data policy on ICES website. All extracts must be acknowledged. For other reproduction requests please contact the General Secretary. This document is the product of an expert group under the auspices of the International Council for the Exploration of the Sea and does not necessarily represent the view of the Council. ISSN number: 2618-1371 I © 2020 International Council for the Exploration of the Sea ICES Scientific Reports Volume 2 | Issue 76 WORKSHOP ON THE DISTRIBUTION AND BYCATCH MANAGEMENT OP- TIONS OF LISTED DEEP-SEA SHARK SPECIES (WKSHARK6) Recommended format for purpose of citation: ICES. 2020. Workshop on the distribution and bycatch management options of listed deep-sea shark species (WKSHARK6). -

251 Part 640—Spiny Lobster Fishery of the Gulf Of
Fishery Conservation and Management Pt. 640 of an application for an ILAP or an ap- Sevengill, Heptranchias perlo peal of NMFS's denial of an initial lim- Sixgill, Hexanchus griseus ited access permit for swordfish. Smalltail, Carcharhinus porosus (12) Falsify information submitted Whale, Rhincodon typus White, Carcharodon carcharias under § 635.46(b) in support of entry of imported swordfish. TABLE 2 OF APPENDIX A TO PART 635± (13) Exceed the incidental catch re- DEEPWATER/OTHER SHARK SPECIES tention limits specified at § 635.24(b). Blotched catshark, Scyliorhinus meadi [64 FR 29135, May 28, 1999, as amended at 64 Broadgill catshark, Apristurus riveri FR 37705, July 13, 1999; 65 FR 42887, July 12, Chain dogfish, Scyliorhinus retifer Deepwater catshark, Apristurus profundorum 2000; 65 FR 47238, Aug. 1, 2000] Dwarf catshark, Scyliorhinus torrei Iceland catshark, Apristurus laurussoni APPENDIX A TO PART 635ÐSPECIES Marbled catshark, Galeus arae TABLES Smallfin catshark, Apristurus parvipinnis TABLE 1 OF APPENDIX A TO PART 635±OCEANIC Bigtooth cookiecutter, Isistius plutodus Blainville's dogfish, Squalus blainvillei SHARKS Bramble shark, Echinorhinus brucus A. Large coastal sharks: Broadband dogfish, Etmopterus gracilispinnis Caribbean lanternshark, Etmopterus hillianus 1. Ridgeback sharks: Cookiecutter shark, Isistius brasiliensis Sandbar, Carcharhinus plumbeus Cuban dogfish, Squalus cubensis Silky, Carcharhinus falciformis Flatnose gulper shark, Deania profundorum Tiger, Galeocerdo cuvieri Fringefin lanternshark, Etmopterus schultzi Great -

Impaginato Quaderno 24
Quaderno di Studi e Notizie di Storia Naturale della Romagna Quad. Studi Nat. Romagna, 24: 19-36, giugno 2007 ISSN 1123-6787 Stefano Marsili PLIOCENE ELASMOBRANCHS IN THE COLLECTION OF THE “MUSEO CIVICO GIUSEPPE SCARABELLI” OF IMOLA (Pisces Chondrichthyes Neoselachii) Abstract Seven elasmobranch species, Carcharias acutissima, Isurus oxyrinchus, I. xiphodon, Carcharodon carcharias, Carcharhinus aff. plumbeus, Carcharhinus sp., Scymnodon cf. ringens, and an indeterminate Myliobatidae, are recognised by the revision of the elasmobranch teeth collection of the “Museo Civico Giuseppe Scarabelli” of Imola (Bologna). The presence of the family Myliobatidae is confi rmed by a fragmentary caudal spine not assigned at generic level. The present study provides a relevant contribution to the understanding of the Pliocene elasmobranch diversity of Romagna region. Riassunto [Elasmobranchi pliocenici nella collezione del “Museo Civico Giuseppe Scarabelli” di Imola] Sette specie di elasmobranchi, Carcharias acutissima, Isurus oxyrinchus, I. xiphodon, Carcharodon carcharias, Carcharhinus aff. plumbeus, Carcharhinus sp., Scymnodon cf. ringens e un Myliobatidae non determinabile, sono stati identifi cati nell’ambito della revisione della collezione ad ittiodontoliti conservata presso il “Museo Civico Giuseppe Scarabelli” di Imola (Bologna). La presenza della famiglia Myliobatidae è confermata sulla base di un piccolo frammento di spina caudale non identifi cabile a livello tassonomico generico. Il presente studio fornisce un ulteriore contributo alla conoscenza della diversità della fauna pliocenica ad elasmobranchi della Romagna. Key words: Teeth, Sharks, Elasmobranch, fi sh fauna, Pliocene, Mediterranean, Romagna, Italy. 19 Introduction Elasmobranchs are poorly represented in the Neogene deposits of Romagna Re- gion. SCARABELLI (1851a, b) fi rst recorded shark teeth from the Miocene sedi- ments cropping out near the Casola Valsenio (Ravenna), and Imola (Bologna) villages. -

And Their Functional, Ecological, and Evolutionary Implications
DePaul University Via Sapientiae College of Science and Health Theses and Dissertations College of Science and Health Spring 6-14-2019 Body Forms in Sharks (Chondrichthyes: Elasmobranchii), and Their Functional, Ecological, and Evolutionary Implications Phillip C. Sternes DePaul University, [email protected] Follow this and additional works at: https://via.library.depaul.edu/csh_etd Part of the Biology Commons Recommended Citation Sternes, Phillip C., "Body Forms in Sharks (Chondrichthyes: Elasmobranchii), and Their Functional, Ecological, and Evolutionary Implications" (2019). College of Science and Health Theses and Dissertations. 327. https://via.library.depaul.edu/csh_etd/327 This Thesis is brought to you for free and open access by the College of Science and Health at Via Sapientiae. It has been accepted for inclusion in College of Science and Health Theses and Dissertations by an authorized administrator of Via Sapientiae. For more information, please contact [email protected]. Body Forms in Sharks (Chondrichthyes: Elasmobranchii), and Their Functional, Ecological, and Evolutionary Implications A Thesis Presented in Partial Fulfilment of the Requirements for the Degree of Master of Science June 2019 By Phillip C. Sternes Department of Biological Sciences College of Science and Health DePaul University Chicago, Illinois Table of Contents Table of Contents.............................................................................................................................ii List of Tables..................................................................................................................................iv -

© Iccat, 2007
A2.2 ICCAT Species Codes APPENDIX 2.2: SPECIES CODES Y ello wfin tuna Codes used to identify the ICCAT tuna and tuna-like species as well as by-catch species Atún blanco Tuna and tuna-like species G e r m o n Numerical Alphabetical Scientific Name English SkipjackFra tunancais EspañolR a b i l 1 BFT Thunnus thynnus Northern bluefin tuna Thon rouge du Nord Atún común (Cimarrón) 2 SBF Thunnus maccoyii Southern bluefin tuna Thon rouge du Sud Atún del Sur 3 YFT Thunnus albacares erocablA T hazard-bâtard L i s t a d o 4 ALB Thunnus alalunga erocablA Plain bonito 5 BET Thunnus obesus Bigeye tuna Thon obèse(=Patudo)P a l o m e t tPatudo e 6 BLF Thunnus atlanticus Blackfin tuna Thon à nageoires noires Atún des aletas negras 7 LTA Euthynnus alletteratus Little tunny(=Atl.black skipjack) Thonine commune BacoretaT a s a r t e 8 SKJ Katsuwonus pelamis WBlack a h o o m arlinoatsiL M akaire noir 9 BON Sarda sarda Atlantic bonito Bonite à dos rayé Bonito del AtlánticoA guja negra P e t o 10 FRI Auxis thazard Frigate tuna Auxide Melva 11 BOP Orcynopsis unicolor 12 WAH Acanthocybium solandri Pez espada 13 SSM Scomberomorus maculatus Atlantic SpanishS w mackerel o r d f i s hTh azard atlantique Carite atlántico 14 KGM Scomberomorus cavalla King mackerel Thazard Ebarr sé p a d o n Carite lucio 15 SAI Istiophorus albicans Atlantic sailfish Voilier de l'Atlantique Pez vela del Atlántico 16 BLM Makaira indica 17 BUM Makaira nigricans Atlantic blue marlin Makaire bleu de l'Atlantique Aguja azul del Atlántico 18 WHM Tetrapturus albidus Atlantic white marlin Makaire blanc de l'Atlantique Aguja blanca del Atlántico 28 19 SWO Xiphias gladius 3 20 SPF Tetrapturus pfluegeri Longbill spearfish Makaire bécune Aguja picuda 284 ICCAT MANUAL, 1st Edition (January 2010) 21 TUN Thunnini sanuT ien sédinohT acn senutA pen 23 YOU gnuoY sanut senueJ sédinoht senutA senevój 24 BIL Istiophoridae Marlins,sailfishes,etc.