LAC.12 Embryology 2019-2020 Dr.Mahdi Alheety
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Te2, Part Iii
TERMINOLOGIA EMBRYOLOGICA Second Edition International Embryological Terminology FIPAT The Federative International Programme for Anatomical Terminology A programme of the International Federation of Associations of Anatomists (IFAA) TE2, PART III Contents Caput V: Organogenesis Chapter 5: Organogenesis (continued) Systema respiratorium Respiratory system Systema urinarium Urinary system Systemata genitalia Genital systems Coeloma Coelom Glandulae endocrinae Endocrine glands Systema cardiovasculare Cardiovascular system Systema lymphoideum Lymphoid system Bibliographic Reference Citation: FIPAT. Terminologia Embryologica. 2nd ed. FIPAT.library.dal.ca. Federative International Programme for Anatomical Terminology, February 2017 Published pending approval by the General Assembly at the next Congress of IFAA (2019) Creative Commons License: The publication of Terminologia Embryologica is under a Creative Commons Attribution-NoDerivatives 4.0 International (CC BY-ND 4.0) license The individual terms in this terminology are within the public domain. Statements about terms being part of this international standard terminology should use the above bibliographic reference to cite this terminology. The unaltered PDF files of this terminology may be freely copied and distributed by users. IFAA member societies are authorized to publish translations of this terminology. Authors of other works that might be considered derivative should write to the Chair of FIPAT for permission to publish a derivative work. Caput V: ORGANOGENESIS Chapter 5: ORGANOGENESIS -

Abnormal Embryonic Lymphatic Vessel Development in Tie1 Hypomorphic Mice Xianghu Qu, Kevin Tompkins, Lorene E
© 2014. Published by The Company of Biologists Ltd | Development (2014) 141, 1417 doi:10.1242/dev.108969 CORRECTION Abnormal embryonic lymphatic vessel development in Tie1 hypomorphic mice Xianghu Qu, Kevin Tompkins, Lorene E. Batts, Mira Puri and H. Scott Baldwin There was an error published in Development 137, 1285-1295. Author name H. Scott Baldwin was incomplete. The correct author list appears above. The authors apologise to readers for this mistake. 1417 RESEARCH ARTICLE 1285 Development 137, 1285-1295 (2010) doi:10.1242/dev.043380 © 2010. Published by The Company of Biologists Ltd Abnormal embryonic lymphatic vessel development in Tie1 hypomorphic mice Xianghu Qu1, Kevin Tompkins1, Lorene E. Batts1, Mira Puri2 and Scott Baldwin1,3,* SUMMARY Tie1 is an endothelial receptor tyrosine kinase that is essential for development and maintenance of the vascular system; however, the role of Tie1 in development of the lymphatic vasculature is unknown. To address this question, we first documented that Tie1 is expressed at the earliest stages of lymphangiogenesis in Prox1-positive venous lymphatic endothelial cell (LEC) progenitors. LEC Tie1 expression is maintained throughout embryonic development and persists in postnatal mice. We then generated two lines of Tie1 mutant mice: a hypomorphic allele, which has reduced expression of Tie1, and a conditional allele. Reduction of Tie1 levels resulted in abnormal lymphatic patterning and in dilated and disorganized lymphatic vessels in all tissues examined and in impaired lymphatic drainage in embryonic skin. Homozygous hypomorphic mice also exhibited abnormally dilated jugular lymphatic vessels due to increased production of Prox1-positive LECs during initial lymphangiogenesis, indicating that Tie1 is required for the early stages of normal lymphangiogenesis. -

CCM2 and CCM3 Proteins Contribute to Vasculogenesis and Angiogenesis in Human Placenta
Histol Histopathol (2009) 24: 1287-1294 Histology and http://www.hh.um.es Histopathology Cellular and Molecular Biology CCM2 and CCM3 proteins contribute to vasculogenesis and angiogenesis in human placenta Gamze Tanriover1, Yasemin Seval1, Leyla Sati1, Murat Gunel2 and Necdet Demir1 1Department of Histology and Embryology, Akdeniz University, School of Medicine, Antalya, Turkey and 2 Department of Neurosurgery, Yale University, School of Medicine, New Haven, CT, USA Summary. Placenta as an ideal model to study Introduction angiogenic mechanisms have been established in previous studies. There are two processes, The placenta is a multifaceted organ that plays a vasculogenesis and angiogenesis, involved in blood critical role in maintaining and protecting the developing vessel formation during placental development. fetus. Normal development and function of the placenta Therefore, blood vessel formation is a crucial issue that requires extensive vasculogenesis and subsequent might cause vascular malformations. One of the vascular angiogenesis, in both maternal and fetal tissues. malformations is cerebral cavernous malformation Vasculogenesis is the formation of the primitive vascular (CCM) in the central nervous system, consisting of network de novo from progenitor cells, and angiogenesis endothelium-lined vascular channels without intervening is identified as the extension of blood vessels from normal brain parenchyma. Three CCM loci have been preexisting vascular structures (Demir et al., 1989, 2006; mapped as Ccm1, Ccm2, Ccm3 genes in CCM. In order Geva et al., 2002; Charnock-Jones et al., 2004). Many to investigate whether CCM proteins participate in blood factors, such as vascular endothelial growth factor vessel formation, we report here the expression patterns (VEGF), angiopoietins (Angpt-1 and -2) and their of CCM2 and CCM3 in developing and term human receptors are involved in the molecular regulation of placenta by means of immunohistochemistry and these diverse developmental steps. -

PDF Published Online: 21 May 2010
Int. J. Dev. Biol. 54: 1033-1043 (2010) DEVELOPMENTALTHE INTERNATIONAL JOURNAL OF doi: 10.1387/ijdb.103105gs BIOLOGY www.intjdevbiol.com Primitive and definitive erythropoiesis in the yolk sac: a bird’s eye view GUOJUN SHENG* Laboratory for Early Embryogenesis, RIKEN Center for Developmental Biology, Kobe, Hyogo, Japan ABSTRACT The yolk sac is the sole niche and source of cells for primitive erythropoiesis from E1 to E5 of chicken development. It is also the main niche and source of cells for early definitive erythropoiesis from E5 to E12. A transition occurs during late embryonic development, after which the bone marrow becomes the major niche and intraembryonically-derived cells the major source. How the yolk sac is involved in these three phases of erythropoiesis is discussed in this review. Prior to the establishment of circulation at E2, specification of primitive erythrocytes is discussed in relation to that of two other cell types formed in the extraembryonic mesoderm, namely the smooth muscle and endothelial cells. Concepts of blood island, hemangioblast and hemogenic endothelium are also discussed. It is concluded that the chick embryo remains a powerful model for studying developmental hematopoiesis and erythropoiesis. KEY WORDS: chicken, primitive erythropoiesis, definitive erythropoiesis, hematopoiesis, yolk sac Introduction 2003; Siatskas and Boyd, 2000), hematological description of different blood lineages (Lucas and Jamroz, 1961), early vascular Studies on chicken hematopoietic development have been morphogenesis (Drake, 2003; Drake et al., 1998; Drake et al., instrumental in the establishment of several key concepts in the 2000) and developmental and adult immune systems (Davison et field, including B lymphocytes, hematopoietic stem cells, al., 2008). -

Problem Is Also Further Confused by the Perplexing Mixture of Cells of Different Origin Brought About by the Early Established Circulation of the Body Fluids
556 ZOOLOGY: C. R. STOCKARD composition. Coralium and Tubipora, for example, are compact forms, with little organic matter; and they are lower in magnesia than the genera with horny, organic axes, such as appear at the eric of the table. It is also noteworthy that the highest proportions of calcium phosphate are commonly found associated with high values for magnesia. 1 Published by permission of the Director of the U. S. Geological Survey. AN EXPERIMENTAL ANALYSIS OF THE ORIGIN AND RELA- TIONSHIP OF BLOOD CORPUSCLES AND THE LINING CELLS OF VESSELS By Charles R. Stockard DEPARTMENT OF ANATOMY, CORNELL UNIVERSITY MEDICAL SCHOOL Preented to the Academy. September 29. 1915 Studies on the origin and development of the cellular elements of the blood and the so-called endothelial cells which line the blood vessels in the normal embryo are peculiarly difficult on account of the important r61e that wandering mesenchyme cells play in these processes. The problem is also further confused by the perplexing mixture of cells of different origin brought about by the early established circulation of the body fluids. The development of no other embryonic tissue is so disturbed by mechanical and physical conditions. A study of living fish embryos with the high power microscope has made it possible to observe the behavior of the wandering cells and to follow them in their development. The disadvantages due to the inter- mixture of cells in the blood current have been overcome by the investi- gation of embryos in which a circulation of the blood is prevented from taking place. When the eggs of the fish, Fundulus heteroclitus, are treated during early developmental stages with weak solutions of alcohol, the resulting embryos in many cases never establish a blood circulation. -

Genetic and Flow Anomalies in Congenital Heart Disease
Published online: 2021-05-10 AIMS Genetics, 3(3): 157-166. DOI: 10.3934/genet.2016.3.157 Received: 01 July 2016 Accepted: 16 August 2016 Published: 23 August 2016 http://www.aimspress.com/journal/Genetics Review Genetic and flow anomalies in congenital heart disease Sandra Rugonyi* Department of Biomedical Engineering, Oregon Health & Science University, 3303 SW Bond Ave. M/C CH13B, Portland, OR 97239, USA * Correspondence: Email: [email protected]; Tel: +1-503-418-9310; Fax: +1-503-418-9311. Abstract: Congenital heart defects are the most common malformations in humans, affecting approximately 1% of newborn babies. While genetic causes of congenital heart disease have been studied, only less than 20% of human cases are clearly linked to genetic anomalies. The cause for the majority of the cases remains unknown. Heart formation is a finely orchestrated developmental process and slight disruptions of it can lead to severe malformations. Dysregulation of developmental processes leading to heart malformations are caused by genetic anomalies but also environmental factors including blood flow. Intra-cardiac blood flow dynamics plays a significant role regulating heart development and perturbations of blood flow lead to congenital heart defects in animal models. Defects that result from hemodynamic alterations recapitulate those observed in human babies, even those due to genetic anomalies and toxic teratogen exposure. Because important cardiac developmental events, such as valve formation and septation, occur under blood flow conditions while the heart is pumping, blood flow regulation of cardiac formation might be a critical factor determining cardiac phenotype. The contribution of flow to cardiac phenotype, however, is frequently ignored. -

Extensive Vasculogenesis, Angiogenesis, and Organogenesis Precede Lethality in Mice Lacking All V Integrins
Cell, Vol. 95, 507–519, November 13, 1998, Copyright ©1998 by Cell Press Extensive Vasculogenesis, Angiogenesis, and Organogenesis Precede Lethality in Mice Lacking All ␣v Integrins Bernhard L. Bader,*‡ Helen Rayburn,* their ligands. Significant expression of ␣v integrins has Denise Crowley,* and Richard O. Hynes*† been noted, in particular, in neural crest cells (Delannet * Howard Hughes Medical Institute et al., 1994), glial cells (Hirsch et al., 1994; Milner and Center for Cancer Research ffrench-Constant, 1994), muscle (Hirsch et al., 1994; Mc- and Department of Biology Donald et al., 1995; Martin and Sanes, 1997), osteoclasts Massachusetts Institute of Technology (Va¨ a¨ na¨ nen and Horton, 1995), epithelia (␣v6; Breuss et Cambridge, Massachusetts 02139 al., 1995; Huang et al., 1996), and blood vessels during development (Brooks et al., 1994a; Drake et al., 1995; Friedlander et al., 1995, 1996) or angiogenesis in re- Summary sponse to tumors (Brooks et al., 1994a, 1994b, 1996, 1998; Varner et al., 1995). ␣v integrins have been implicated in many develop- Among the ligands for various ␣v integrins is fibro- mental processes and are therapeutic targets for inhi- nectin (FN), and results on mouse embryos lacking FN bition of angiogenesis and osteoporosis. Surprisingly, or FN receptor integrins suggest that ␣v integrins might ablation of the gene for the ␣v integrin subunit, elimi- be important receptors for FN during early development. nating all five ␣v integrins, although causing lethality, FN-null embryos fail to form notochord or somites allows considerable development and organogenesis (George et al., 1993; Georges-Labouesse et al., 1996), including, most notably, extensive vasculogenesis and whereas embryos null for either (Yang et al., 1993, 1995) angiogenesis. -

Copyright by Steven A. Vokes 2002 the Dissertation Committee for Steven Alexander Vokes Certifies That This Is the Approved Version of the Following Dissertation
Copyright by Steven A. Vokes 2002 The Dissertation Committee for Steven Alexander Vokes Certifies that this is the approved version of the following dissertation: The Role of Endoderm in Vascular Patterning Committee: Janice A. Fischer, Supervisor Paul A. Krieg , Co-Supervisor Alan M. Lloyd Arlen W. Johnson S. Martin Shankland R. Adron Harris The Role of Endoderm in Vascular Patterning by Steven Alexander Vokes, B.A. Dissertation Presented to the Faculty of the Graduate School of The University of Texas at Austin in Partial Fulfillment of the Requirements for the Degree of Doctor of Philosophy The University of Texas at Austin December, 2002 Dedication This work, symbolic of my higher education, is dedicated to my parents Carol and Emmett Vokes, who played such an integral role in its foundations. Acknowledgements I have been extremely fortunate to have excellent mentoring during my time in graduate school. I thank Paul Krieg for his suggestions, enthusiasm, encouragement and friendship during this learning process. He has taught me how to think (and write) critically and insightfully about science. I am also grateful to Amy Cheng Vollmer, my undergraduate mentor. She introduced me to the joys of scientific research, and continues to give me excellent advice whenever I need it most. I thank Peter Vize, whose conversations led to the first experiments described within. In daily interactions, I have benefited from a caste of talented co-workers and advisors. These include Craig Newman, Wendy Gerber, Ondine Cleaver, Tom Carroll, Eric Small, Rob Garriock, Martha Joe, Parker Antin, Ray Runyan, Jean Wilson, Carol Gregorio and all past and present members of the Krieg lab. -

The Evolving Cardiac Lymphatic Vasculature in Development, Repair and Regeneration
REVIEWS The evolving cardiac lymphatic vasculature in development, repair and regeneration Konstantinos Klaourakis 1,2, Joaquim M. Vieira 1,2,3 ✉ and Paul R. Riley 1,2,3 ✉ Abstract | The lymphatic vasculature has an essential role in maintaining normal fluid balance in tissues and modulating the inflammatory response to injury or pathogens. Disruption of normal development or function of lymphatic vessels can have severe consequences. In the heart, reduced lymphatic function can lead to myocardial oedema and persistent inflammation. Macrophages, which are phagocytic cells of the innate immune system, contribute to cardiac development and to fibrotic repair and regeneration of cardiac tissue after myocardial infarction. In this Review, we discuss the cardiac lymphatic vasculature with a focus on developments over the past 5 years arising from the study of mammalian and zebrafish model organisms. In addition, we examine the interplay between the cardiac lymphatics and macrophages during fibrotic repair and regeneration after myocardial infarction. Finally, we discuss the therapeutic potential of targeting the cardiac lymphatic network to regulate immune cell content and alleviate inflammation in patients with ischaemic heart disease. The circulatory system of vertebrates is composed of two after MI. In this Review, we summarize the current complementary vasculatures, the blood and lymphatic knowledge on the development, structure and function vascular systems1. The blood vasculature is a closed sys- of the cardiac lymphatic vasculature, with an emphasis tem responsible for transporting gases, fluids, nutrients, on breakthroughs over the past 5 years in the study of metabolites and cells to the tissues2. This extravasation of cardiac lymphatic heterogeneity in mice and zebrafish. -

Glossary of Key Terms and Concepts - Chapter 8
Glossary of Key Terms and Concepts - Chapter 8 Angioblasts - These "vessel-forming cells" may arise from any kind of mesoderm except prechordal plate mesoderm. Angioblastic cords - Angiocysts coalesce to form short blind-ended angioblastic cords. Angioblastic plexuses - Angioblastic cords coalesce to form complex interconnected vascular networks or plexuses. Angiocysts - These vesicles are formed by angioblasts during the process of vasculogenesis. Angiogenesis - This is the mechanism whereby preexisting vessels lengthen or branch by sprouting. Aortic arches - These vessels have been modified in humans to form the great vessels of the thorax (also see Ch. 7). Axis arteries - These central arteries of the limbs are derived from the 7th intersegmental arteries (upper limb) and 5th lumbar intersegmental arteries (lower limb). Blood islands - Blood islands are cysts of angioblasts containing hemoblasts. These coalesce to form blood vessels in the yolk sac and also form the coronary vasculature. Branchial arches - These are the gill bars of fish. Homologous structures of humans are more appropriately named "pharyngeal" arches. Cardinal system of veins - These veins drain the head and neck and body wall and extremities of the embryo. Anterior cardinals drain the head and neck and the trunk and lower extremities are drained by paired posterior cardinals. The posterior cardinal veins are replaced by subcardinal and supracardinal veins during the second month. Coronary vessels - These vessels of the heart form from epicardium as subepicardial plexuses fuse with sprouts of the aorta and coronary sinus to form the coronary arteries and coronary veins respectively. Endothelial cells - These cells arise from angioblasts to form the initial vascular network. -

VEGFR-3 in Angiogenesis and Lymphangiogenesis
VEGFR-3 in Angiogenesis and Lymphangiogenesis Lotta Jussila Molecular/Cancer Biology Laboratory Haartman Institute and Helsinki University Central Hospital Biomedicum Helsinki University of Helsinki Finland Academic dissertation To be publicly discussed, with the permission of the Medical Faculty of the University of Helsinki, in the lecture hall 3 of the Biomedicum Helsinki, Haartmaninkatu 8, Helsinki, on December 14th, 2001 at 12 o´clock noon. Helsinki, 2001 Supervised by Dr. Kari Alitalo Molecular/Cancer Biology Laboratory University of Helsinki Reviewed by Dr. Ulf Eriksson Ludwig Institute for Cancer Research Karolinska Institute and Dr. Hannu Sariola Institute of Biomedicine University of Helsinki Opponent Dr. Christer Betsholtz Department of Medical Biochemistry University of Göteborg ISBN 952-91-4175-0 (nid.) ISBN 952-10-0241-7 (pdf) Multiprint Oy Helsinki VEGFR-3 in Angiogenesis and Lymphangiogenesis 1 Contents Contents............................................................................................................. 1 Abbreviations ....................................................................................................... 2 List of Original Publications ...................................................................................... 3 Abstract ............................................................................................................. 4 Review of the literature .......................................................................................... 5 Blood vessel development ........................................................................... -
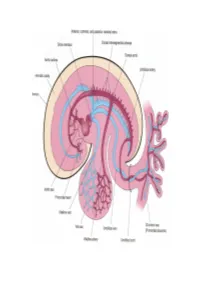
Development of HEART 4-VEINS
Development of brachiocephalic veins 1. Right brachiocephalic vein is formed by cranial part of right anterior cardinal vein and 2. Left brachiocephalic is formed by cranial part of left anterior cardinal vein and the interant.cardinal anastomosis. Development of superior vena cava 1. The part up to the opening of vena azygos develops from caudal part of right ant.cardinal vein and 2. The part below the opening (intrapericardial part) is formed by the right common cardinal vein. Development of azygos and hemiazygos veins A. 1. Vena azygos develops from right azygos line vein and 2. The arch of vena azygos is formed by the cranial end of right postcardinal vein. B. Hemiazygos veins are formed by the left azygos line vein. Development of Inferior vena cava Inferior vena cava is formed, from below upwards by: 1. Begins by the union of the two common iliac veins (postcardinal veins), 2. Right supracardinal, 3. Right supra-subcardinal anastomosis, 4. Right subcardinal, 5. New formation (hepatic segment) and 6. Hepatocardiac channel (terminal part of right vitelline vein). Congenital anomalies • Double inferior vena cava • Absence • Left SVC • Double SVC DEVELOPMENT OF PORTAL VEIN 1. The portal vein is formed behind the neck of pancreas by the union of superior mesentric and splenic vein to the left vitelline vein. 2. The part of the portal vein which is behind the Ist part of duodenum is formed by middle dorsal transverse anastomosis. 3. Part of portal vein which is in the free margin of lesser omentum is formed by cranial or distal part of right vitelline vein.