The Migraine Attack As a Homeostatic, Neuroprotective Response to Brain Oxidative Stress: Preliminary Evidence for a Theory
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Ayahuasca: Spiritual Pharmacology & Drug Interactions
Ayahuasca: Spiritual Pharmacology & Drug Interactions BENJAMIN MALCOLM, PHARMD, MPH [email protected] MARCH 28 TH 2017 AWARE PROJECT Can Science be Spiritual? “Science is not only compatible with spirituality; it is a profound source of spirituality. When we recognize our place in an immensity of light years and in the passage of ages, when we grasp the intricacy, beauty and subtlety of life, then that soaring feeling, that sense of elation and humility combined, is surely spiritual. The notion that science and spirituality are somehow mutually exclusive does a disservice to both.” – Carl Sagan Disclosures & Disclaimers No conflicts of interest to disclose – I don’t get paid by pharma and have no potential to profit directly from ayahuasca This presentation is for information purposes only, none of the information presented should be used in replacement of medical advice or be considered medical advice This presentation is not an endorsement of illicit activity Presentation Outline & Objectives Describe what is known regarding ayahuasca’s pharmacology Outline adverse food and drug combinations with ayahuasca as well as strategies for risk management Provide an overview of spiritual pharmacology and current clinical data supporting potential of ayahuasca for treatment of mental illness Pharmacology Terms Drug ◦ Term used synonymously with substance or medicine in this presentation and in pharmacology ◦ No offense intended if I call your medicine or madre a drug! Bioavailability ◦ The amount of a drug that enters the body and is able to have an active effect ◦ Route specific: bioavailability is different between oral, intranasal, inhalation (smoked), and injected routes of administration (IV, IM, SC) Half-life (T ½) ◦ The amount of time it takes the body to metabolize/eliminate 50% of a drug ◦ E.g. -

Altered Inflammatory Mediators in Fibromyalgia
Cur gy: ren lo t o R t e a s e m a u r c e h h García et al., Rheumatology (Sunnyvale) 2017, 7:2 R Rheumatology: Current Research DOI: 10.4172/2161-1149.1000215 ISSN: 2161-1149 Review Article Open Access Altered Inflammatory Mediators in Fibromyalgia Juan José García1*, Julián Carvajal-Gil2, Aurora Herrero-Olea2 and Rafael Gómez-Galán2 1Department of Physiology, University Centre of Mérida, University of Extremadura, Mérida, Spain 2Department of Nursing, University Centre of Mérida, University of Extremadura, Mérida, Spain *Corresponding author: Dr. Juan José García, Department of Physiology, University Centre of Mérida, University of Extremadura, Mérida, Spain,Tel: 00 34 924289300, Ext: 86118; E-mail: [email protected] Received date: February 07, 2017; Accepted date: April 08, 2017; Published date: April 15, 2017 Copyright: ©2017 García JJ, et al. This is an open access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited. Abstract Fibromyalgia (FM) is a syndrome characterized by widespread chronic pain, tenderness, stiffness, fatigue, and sleep and mood disturbances. Current evidences suggest that inflammatory mediators may have an important role in the pathogenesis of fibromyalgia. Every day new evidences emerging of the role of the immune system and the inflammatory process in the pathophysiology of this disease. Thus, the aim of this work has been review that altered inductors, inflammatory mediators and effectors have been reported in fibromyalgia, and its relationship to disease’s symptoms. If in fibromyalgia underlies widespread disruption of most characteristic components on the inflammatory ´s process (endogenous inductors, mediators and effectors) is logical to think that fibromyalgia is producing a sustained inflammatory response in time, unregulated, responsible for the disease´s symptoms. -

PMA-Sio2: Heteropolyacid Catalysis for Michael Addition-Convenient Route to Substituted-3-Indoles
Available online at www.derpharmachemica.com ISSN 0975-413X Der Pharma Chemica, 2017, 9(13):112-117 CODEN (USA): PCHHAX (http://www.derpharmachemica.com/archive.html) PMA-SiO2: Heteropolyacid Catalysis for Michael Addition-Convenient Route to Substituted-3-Indoles Vijay Kumar Pasala* Deapartment of Chemistry, Osmania University, Hyderabad-500007, Telangana, India ABSTRACT Synthesis of 3-substituted indoles in a hassel-free, ecofriendly manner by treating indoles with α, β-unsaturated carbonyl or nitro compounds under acidic conditions to give good to excellent yields with shorter reaction durations are described. The catalyst is recyclable for three to four times without great loss in the activity. Keywords: Phosphomolybdic Acid (PMA), Substituted indoles, α, β-unsaturated carbonyl compounds, α, β-unsaturated nitro compounds, Michael addition INTRODUCTION Heterocyclic chemistry is one of the quintessential branches of organic synthesis beaconing towards new scaffolds with medicinal values, new methodologies to the existing active principles etc. One among such skeletons is indole. It is perhaps the most common heterocycles in chemistry and its derivatives are obtained from coal pitch, variety of plants or by the bacterial decay of tryptophan in the intestine [1]. Indole derivatives serve as signaling chemicals in plants and animals, as nucleus building blocks (serotonin [2] (A) a crucial neurotransmitter in the central nervous system [3]), as antibacterial [4], antiviral [5], protein kinase inhibitors [6], anticancer agents [7], entheogens (psilocybin (B) causes perceptional changes), hormones (melatonin (C) regulates sleep and wakefulness), antidepressants (roxindole (D), bufotenin (E)), sumatriptan (F) for the treatment of migraine and ondansetron (G) for the suppression of nausea and vastly found in natural products such as alkaloids (Corynanthe, Iboga, and Aspidosperma alkaloids) [8-10], indigoids etc., which originate, either fully or partly, from bio-oxidation of indoles [11]. -

Pharmacokinetics, Pharmacodynamics and Drug
pharmaceutics Review Pharmacokinetics, Pharmacodynamics and Drug–Drug Interactions of New Anti-Migraine Drugs—Lasmiditan, Gepants, and Calcitonin-Gene-Related Peptide (CGRP) Receptor Monoclonal Antibodies Danuta Szkutnik-Fiedler Department of Clinical Pharmacy and Biopharmacy, Pozna´nUniversity of Medical Sciences, Sw.´ Marii Magdaleny 14 St., 61-861 Pozna´n,Poland; [email protected] Received: 28 October 2020; Accepted: 30 November 2020; Published: 3 December 2020 Abstract: In the last few years, there have been significant advances in migraine management and prevention. Lasmiditan, ubrogepant, rimegepant and monoclonal antibodies (erenumab, fremanezumab, galcanezumab, and eptinezumab) are new drugs that were launched on the US pharmaceutical market; some of them also in Europe. This publication reviews the available worldwide references on the safety of these anti-migraine drugs with a focus on the possible drug–drug (DDI) or drug–food interactions. As is known, bioavailability of a drug and, hence, its pharmacological efficacy depend on its pharmacokinetics and pharmacodynamics, which may be altered by drug interactions. This paper discusses the interactions of gepants and lasmiditan with, i.a., serotonergic drugs, CYP3A4 inhibitors, and inducers or breast cancer resistant protein (BCRP) and P-glycoprotein (P-gp) inhibitors. In the case of monoclonal antibodies, the issue of pharmacodynamic interactions related to the modulation of the immune system functions was addressed. It also focuses on the effect of monoclonal antibodies on expression of class Fc gamma receptors (FcγR). Keywords: migraine; lasmiditan; gepants; monoclonal antibodies; drug–drug interactions 1. Introduction Migraine is a chronic neurological disorder characterized by a repetitive, usually unilateral, pulsating headache with attacks typically lasting from 4 to 72 h. -

Sustained Sensitizing Effects of Tumor Necrosis Factor Alpha on Sensory Nerves in Lung and Airways Ruei-Lung Lin University of Kentucky, [email protected]
University of Kentucky UKnowledge Physiology Faculty Publications Physiology 12-2017 Sustained Sensitizing Effects of Tumor Necrosis Factor Alpha on Sensory Nerves in Lung and Airways Ruei-Lung Lin University of Kentucky, [email protected] Qihai Gu Mercer University Mehdi Khosravi University of Kentucky, [email protected] Lu-Yuan Lee University of Kentucky, [email protected] Right click to open a feedback form in a new tab to let us know how this document benefits oy u. Follow this and additional works at: https://uknowledge.uky.edu/physiology_facpub Part of the Pharmacology Commons, Physiology Commons, Pulmonology Commons, and the Therapeutics Commons Repository Citation Lin, Ruei-Lung; Gu, Qihai; Khosravi, Mehdi; and Lee, Lu-Yuan, "Sustained Sensitizing Effects of Tumor Necrosis Factor Alpha on Sensory Nerves in Lung and Airways" (2017). Physiology Faculty Publications. 139. https://uknowledge.uky.edu/physiology_facpub/139 This Article is brought to you for free and open access by the Physiology at UKnowledge. It has been accepted for inclusion in Physiology Faculty Publications by an authorized administrator of UKnowledge. For more information, please contact [email protected]. Sustained Sensitizing Effects of Tumor Necrosis Factor Alpha on Sensory Nerves in Lung and Airways Notes/Citation Information Published in Pulmonary Pharmacology & Therapeutics, v. 47, p. 29-37. © 2017 Elsevier Ltd. All rights reserved. This manuscript version is made available under the CC‐BY‐NC‐ND 4.0 license https://creativecommons.org/licenses/by-nc-nd/4.0/. The document available for download is the author's post-peer-review final draft of the ra ticle. Digital Object Identifier (DOI) https://doi.org/10.1016/j.pupt.2017.06.001 This article is available at UKnowledge: https://uknowledge.uky.edu/physiology_facpub/139 HHS Public Access Author manuscript Author ManuscriptAuthor Manuscript Author Pulm Pharmacol Manuscript Author Ther. -

Migraine: Current Concepts and Emerging Therapies
Vascular Pharmacology 43 (2005) 176 – 187 www.elsevier.com/locate/vph Migraine: Current concepts and emerging therapies D.K. Arulmozhi a,b,*, A. Veeranjaneyulu a, S.L. Bodhankar b aNew Chemical Entity Research, Lupin Research Park, Village Nande, Taluk Mulshi, Pune 411 042, Maharashtra, India bDepartment of Pharmacology, Bharati Vidyapeeth, Poona College of Pharmacy, Pune 411 038, Maharashtra, India Received 23 April 2005; received in revised form 17 June 2005; accepted 11 July 2005 Abstract Migraine is a recurrent incapacitating neurovascular disorder characterized by attacks of debilitating pain associated with photophobia, phonophobia, nausea and vomiting. Migraine affects a substantial fraction of world population and is a major cause of disability in the work place. Though the pathophysiology of migraine is still unclear three major theories proposed with regard to the mechanisms of migraine are vascular (due to cerebral vasodilatation), neurological (abnormal neurological firing which causes the spreading depression and migraine) and neurogenic dural inflammation (release of inflammatory neuropeptides). The modern understanding of the pathogenesis of migraine is based on the concept that it is a neurovascular disorder. The drugs used in the treatment of migraine either abolish the acute migraine headache or aim its prevention. The last decade has witnessed the advent of Sumatriptan and the Ftriptan_ class of 5-HT1B/1D receptor agonists which have well established efficacy in treating migraine. Currently prophylactic treatments for migraine include calcium channel blockers, 5-HT2 receptor antagonists, beta adrenoceptor blockers and g-amino butyric acid (GABA) agonists. Unfortunately, many of these treatments are non specific and not always effective. Despite such progress, in view of the complexity of the etiology of migraine, it still remains undiagnosed and available therapies are underused. -

Triptans Step Therapy/Quantity Limit Criteria
Triptans Step Therapy/Quantity Limit Criteria Program may be implemented with the following options 1) step therapy 2) quantity limits or 3) step therapy with quantity limits For Blue Cross and Blue Shield of Illinois Option 1 (step therapy only) will apply. Brand Generic Dosage Form Amerge® naratriptan tablets Axert® almotriptan tablets Frova® frovatriptan tablets Imitrex® sumatriptan injection*, nasal spray, tablets* Maxalt® rizatriptan tablets Maxalt-MLT® rizatriptan tablets Relpax® eletriptan tablets Treximet™ sumatriptan and naproxen tablets Zomig® zolmitriptan tablets, nasal spray Zomig-ZMT® zolmitriptan tablets * generic available and included as target agent in quantity limit edit FDA APPROVED INDICATIONS1-7 The following information is taken from individual drug prescribing information and is provided here as background information only. Not all FDA-approved indications may be considered medically necessary. All criteria are found in the section “Prior Authorization Criteria for Approval.” Amerge® Tablets1, Axert® Tablets2, Frova® Tablets3,Imitrex® injection4, Imitrex Nasal Spray5, Imitrex Tablets6, Maxalt® Tablets7, Maxalt-MLT® Tablets7, Relpax® Tablets8, Treximet™ Tablets9, Zomig® Tablets10, Zomig-ZMT® Tablets10, and Zomig® Nasal Spray11 Amerge (naratriptan), Axert (almotriptan), Frova (frovatriptan), Imitrex (sumatriptan), Maxalt (rizatriptan), Maxalt-MLT (rizatriptan orally disintegrating), Relpax (eletriptan), Treximet (sumatriptan/naproxen), Zomig (zolmitriptan), and Zomig-ZMT (zolmitriptan orally disintegrating) tablets, and Imitrex (sumatriptan) and Zomig (zolmitriptan) nasal spray are all indicated for the acute treatment of migraine attacks with or without aura in adults. They are not intended for the prophylactic therapy of migraine or for use in the management of hemiplegic or basilar migraine. Safety and effectiveness of all of these products have not been established for cluster headache, which is present in an older, predominantly male population. -

European Patent Office of Opposition to That Patent, in Accordance with the Implementing Regulations
(19) TZZ _T (11) EP 2 666 772 B1 (12) EUROPEAN PATENT SPECIFICATION (45) Date of publication and mention (51) Int Cl.: of the grant of the patent: C07D 409/04 (2006.01) C07D 209/16 (2006.01) 01.03.2017 Bulletin 2017/09 C07D 209/18 (2006.01) C07D 209/38 (2006.01) (21) Application number: 13173141.6 (22) Date of filing: 12.12.2003 (54) Synthesis of amines and intermediates for the synthesis thereof Synthese von Aminen und Zwischenverbindungen für die Synthese davon Synthèse d’amines et intermédiaires pour leur synthèse (84) Designated Contracting States: • DEMERSON C A ET AL: "ETODOLIC ACID AND AT BE BG CH CY CZ DE DK EE ES FI FR GB GR RELATED COMPOUNDS CHEMISTRY AND HU IE IT LI LU MC NL PT RO SE SI SK TR ANTIINFLAMMATORY ACTIONS OF SOME POTENT DI-AND TRISUBSTITUTED (30) Priority: 20.12.2002 EP 02406128 1,3,4,9-TETRAHYDROPYRANO not 3,4-B 3/4 INDOLE-I-ACETIC ACIDS", JOURNAL OF (43) Date of publication of application: MEDICINALCHEMISTRY, AMERICAN CHEMICAL 27.11.2013 Bulletin 2013/48 SOCIETY, vol. 19, no. 3, 1 March 1976 (1976-03-01), pages391-395, XP000940626, ISSN: (62) Document number(s) of the earlier application(s) in 0022-2623, DOI: 10.1021/JM00225A010 accordance with Art. 76 EPC: • HARUYASU ET ALL: "Possible metabolic 03799560.2 / 1 572 647 Intermediatesfrom IAA to beta-acid in Rice Bran", AGR. BIOL. CHEM., vol. 40, no. 12, 1976, pages (73) Proprietor: BASF SE 2465-2470, XP002714693, 67056 Ludwigshafen am Rhein (DE) • SOLL R M ET AL: "Multigram preparation of 1,8-diethyl-7-hydroxy-1,3,4,9-tetrahydropy (72) Inventors: rano[3,4-b]indole-1-acetic acid, a phenolic • Berens, Ulrich metabolite of the analgesic and antiinflammatory 79589 Binzen (DE) agent etodolac", THE JOURNAL OF ORGANIC • Dosenbach, Oliver CHEMISTRY, AMERICAN CHEMICAL SOCIETY 79415 Bad Bellingen (DE) [NOT]ETC. -

Patient Information Leaflet
Package Leaflet: Information for the user Sumatriptan 50 mg film-coated tablets Sumatriptan 100 mg film-coated tablets sumatriptan Read all of this leaflet carefully before you start using this medicine because it contains important information for you. • Keep this leaflet. You may need to read it again. • If you have any further questions, ask your doctor or pharmacist. • This medicine has been prescribed for you only. Do not pass it on to others. It may harm them, even if their signs of illness are the same as yours. • If you get any side effects, talk to your doctor or pharmacist. This includes any possible side effects not listed in this leaflet. See section 4. What is in this leaflet 1. What Sumatriptan tablet is and what it is used for 2. What you need to know before you use Sumatriptan tablet 3. How to use Sumatriptan tablet 4. Possible side effects 5. How to store Sumatriptan tablet 6. Contents of the pack and other information 1. What Sumatriptan tablet is and what it is used for Sumatriptan belongs to a group of medicines called serotonin receptor (5-HT1) agonists. Migraine headaches are thought to result from the dilatation of blood vessels. Sumatriptan constricts these blood vessels, thus relieving the migraine headache. Sumatriptan tablet is used to treat migraine attacks with or without aura (aura is a premonition usually connected with flashes of light, serrated images, stars or waves). 2. What you need to know before you use Sumatriptan tablet Do not use Sumatriptan tablet: - if you are allergic to sumatriptan or any of the other ingredients of this medicine (listed in section 6). -
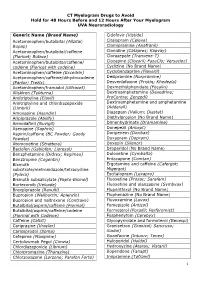
CT Myelogram Drugs to Avoid Hold for 48 Hours Before and 12 Hours After Your Myelogram UVA Neuroradiology
CT Myelogram Drugs to Avoid Hold for 48 Hours Before and 12 Hours After Your Myelogram UVA Neuroradiology Generic Name (Brand Name) Cidofovir (Vistide) Acetaminophen/butalbital (Allzital; Citalopram (Celexa) Bupap) Clomipramine (Anafranil) Acetaminophen/butalbital/caffeine Clonidine (Catapres; Kapvay) (Fioricet; Butace) Clorazepate (Tranxene-T) Acetaminophen/butalbital/caffeine/ Clozapine (Clozaril; FazaClo; Versacloz) codeine (Fioricet with codeine) Cyclizine (No Brand Name) Acetaminophen/caffeine (Excedrin) Cyclobenzaprine (Flexeril) Acetaminophen/caffeine/dihydrocodeine Desipramine (Norpramine) (Panlor; Trezix) Desvenlafaxine (Pristiq; Khedezla) Acetaminophen/tramadol (Ultracet) Dexmethylphenidate (Focalin) Aliskiren (Tekturna) Dextroamphetamine (Dexedrine; Amitriptyline (Elavil) ProCentra; Zenzedi) Amitriptyline and chlordiazepoxide Dextroamphetamine and amphetamine (Limbril) (Adderall) Amoxapine (Asendin) Diazepam (Valium; Diastat) Aripiprazole (Abilify) Diethylpropion (No Brand Name) Armodafinil (Nuvigil) Dimenhydrinate (Dramamine) Asenapine (Saphris) Donepezil (Aricept) Aspirin/caffeine (BC Powder; Goody Doripenem (Doribax) Powder) Doxapram (Dopram) Atomoxetine (Strattera) Doxepin (Silenor) Baclofen (Gablofen; Lioresal) Droperidol (No Brand Name) Benzphetamine (Didrex; Regimex) Duloxetine (Cymbalta) Benztropine (Cogentin) Entacapone (Comtan) Bismuth Ergotamine and caffeine (Cafergot; subcitrate/metronidazole/tetracycline Migergot) (Pylera) Escitalopram (Lexapro) Bismuth subsalicylate (Pepto-Bismol) Fluoxetine (Prozac; Sarafem) -

Actions of the 5-Hydroxytryptamine 1 Receptor Agonist Sumatriptan
36 Gut 1998;42:36–41 Actions of the 5-hydroxytryptamine 1 receptor agonist sumatriptan on interdigestive Gut: first published as 10.1136/gut.42.1.36 on 1 January 1998. Downloaded from gastrointestinal motility in man J Tack, B Coulie, A Wilmer, T Peeters, J Janssens Abstract Evidence has accumulated that enteric neu- Background—Pharmacological studies of ronal 5-hydroxytryptamine (5HT) receptors the enteric nervous system have shown the are involved in regulation of the MMC in ani- presence of several subtypes of 5-hydroxy- mals and in man.3–7 In vitro studies in the tryptamine (5HT) receptor, which might guinea pig showed that 5HT4 receptors, 5HT3 be involved in control of the migrating receptors, 5HT1A receptors, and 5HT1P recep- motor complex. tors are present on myenteric neurones in the Aims—To study the eVect of sumatriptan, stomach and in the small intestine.8–11 Using an agonist of enteric neuronal 5HT1P the selective 5HT3 receptor antagonist on- receptors, on interdigestive motility in dansetron, we showed that 5HT3 receptors are man. involved in the initiation of gastric phase 3 12 Subjects and methods—In 12 healthy sub- motor activity in man. Lack of suitable jects, interdigestive motility was recorded ligands precluded a similar study for 5HT1 manometrically in the upper gastrointes- receptors and for 5HT4 receptors. Recently, tinal tract. In seven subjects blood sam- however, we showed that sumatriptan, a 5HT1 ples were drawn every 15 minutes for receptor agonist which is used in the treatment 13 radioimmunoassay of motilin and soma- of migraine in man, is an agonist of 5HT1P tostatin. -

Infection-Associated Mechanisms of Neuro-Inflammation and Neuro
International Journal of Molecular Sciences Review Infection-Associated Mechanisms of Neuro-Inflammation and Neuro-Immune Crosstalk in Chronic Respiratory Diseases Belinda Camp, Sabine Stegemann-Koniszewski *,† and Jens Schreiber † Experimental Pneumonology, Department of Pneumonology, University Hospital Magdeburg, Health Campus Immunology, Infectiology and Inflammation (GC-I3), Otto-von-Guericke University Magdeburg, 39120 Magdeburg, Germany; [email protected] (B.C.); [email protected] (J.S.) * Correspondence: [email protected] † These authors contributed equally to this work. Abstract: Chronic obstructive airway diseases are characterized by airflow obstruction and airflow limitation as well as chronic airway inflammation. Especially bronchial asthma and chronic obstruc- tive pulmonary disease (COPD) cause considerable morbidity and mortality worldwide, can be difficult to treat, and ultimately lack cures. While there are substantial knowledge gaps with respect to disease pathophysiology, our awareness of the role of neurological and neuro-immunological processes in the development of symptoms, the progression, and the outcome of these chronic obstructive respiratory diseases, is growing. Likewise, the role of pathogenic and colonizing mi- croorganisms of the respiratory tract in the development and manifestation of asthma and COPD is increasingly appreciated. However, their role remains poorly understood with respect to the underly- ing mechanisms. Common bacteria and viruses causing respiratory infections and exacerbations of chronic obstructive respiratory diseases have also been implicated to affect the local neuro-immune Citation: Camp, B.; crosstalk. In this review, we provide an overview of previously described neuro-immune interactions Stegemann-Koniszewski, S.; in asthma, COPD, and respiratory infections that support the hypothesis of a neuro-immunological Schreiber, J.