(Coregonus Lavaretus L.) I Isteren
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Fiskesamfunnet I Storsjøen I Åmot Og Rendalen Kommuner
388 Fiskesamfunnet i Storsjøen i Åmot og Rendalen kommuner Betydningen av reguleringsinngrep, endret beskatning og avbøtende tiltak Jon Museth Odd Terje Sandlund Stein I. Johnsen Sigurd Rognerud Randi Saksgård NINAs publikasjoner NINA Rapport Dette er en ny, elektronisk serie fra 2005 som erstatter de tidligere seriene NINA Fagrapport, NINA Oppdragsmelding og NINA Project Report. Normalt er dette NINAs rapportering til oppdragsgiver etter gjennomført forsknings-, overvåkings- eller utredningsarbeid. I tillegg vil serien favne mye av instituttets øvrige rapportering, for eksempel fra seminarer og konferanser, resultater av eget forsk- nings- og utredningsarbeid og litteraturstudier. NINA Rapport kan også utgis på annet språk når det er hensiktsmessig. NINA Temahefte Som navnet angir behandler temaheftene spesielle emner. Heftene utarbeides etter behov og seri- en favner svært vidt; fra systematiske bestemmelsesnøkler til informasjon om viktige problemstil- linger i samfunnet. NINA Temahefte gis vanligvis en populærvitenskapelig form med mer vekt på illustrasjoner enn NINA Rapport. NINA Fakta Faktaarkene har som mål å gjøre NINAs forskningsresultater raskt og enkelt tilgjengelig for et større publikum. De sendes til presse, ideelle organisasjoner, naturforvaltningen på ulike nivå, politikere og andre spesielt interesserte. Faktaarkene gir en kort framstilling av noen av våre viktigste forsk- ningstema. Annen publisering I tillegg til rapporteringen i NINAs egne serier publiserer instituttets ansatte en stor del av sine vi- tenskapelige resultater i internasjonale journaler, populærfaglige bøker og tidsskrifter. Norsk institutt for naturforskning Fiskesamfunnet i Storsjøen i Åmot og Rendalen kommuner Betydningen av reguleringsinngrep, endret beskatning og avbøtende tiltak Jon Museth Odd Terje Sandlund Stein I. Johnsen Sigurd Rognerud Randi Saksgård NINA Rapport 388 Museth, J., Sandlund, O. -

Kvikksølv I Storørret Og -Røye I Norske Innsjøer, 2000–2001 4502-2002 Mars 2001
RAPPORT LNR 4502-02 Kvikks¿lv i stor¿rret og -r¿ye i norske innsj¿er, 2000Ð2001 4.5 4 3.5 3 2.5 2 1.5 1 omsetningsgrense kvikksølvkonsentrasjon, mg Hg/kg 0.5 0 10 20 30 40 50 60 70 80 90 100 lengde, cm ©Eirik Fjeld Norsk institutt for vannforskning RAPPORT Hovedkontor Sørlandsavdelingen Østlandsavdelingen Vestlandsavdelingen Akvaplan-niva Postboks 173, Kjelsås Televeien 3 Sandvikaveien 41 Nordnesboder 5 0411 Oslo 4979 Grimstad 2312 Otestad 5008 Bergen 9296 Tromsø Telefon (47) 22 18 51 00 Telefon (47) 37 29 50 55 Telefon (47) 67 57 64 00 Telefon (47) 55 30 22 50 Telefon (47) 77 75 03 00 Telefax (47) 22 18 52 00 Telefax (47) 37 04 45 13 Telefax (47) 62 57 66 53 Telefax (47) 55 30 22 51 Telefax (47) 77 75 03 01 Internet: www.niva.no Tittel Løpenr. (for bestilling) Dato Kvikksølv i storørret og -røye i norske innsjøer, 2000–2001 4502-2002 mars 2001 Prosjektnr. Undernr. Sider Pris O-20120 42 Forfattere Fagområde Distribusjon Eirik Fjeld og Sigurd Rognerud Miljøgifter Fri Geografisk område Trykket Norge NIVA Oppdragsgiver(e) Oppdragsreferanse Statens forurensningstilsyn (SFT) Per Erik Iversen Statens næringsmiddeltilsyn (SNT) Marie Wiborg Sammendrag Det rapporteres her en nasjonal undersøkelser over konsentrasjonene av kvikksølv i storørret og stor røye, med hoved- vekt på ørret. Materialet er i hovedsak innsamlet i 2000–2001, men komplementerende data fra tidligere undersøkel- ser er også inkludert. Analysen av ørretmaterialet viste at kvikksølvnivået steg med fiskens størrelse. Ved en lengde på knapt 60 cm (ca. 2,5 kg) var det 50% sannsynlighet for at fisken hadde en kvikksølvkonsentrasjon større enn 0,5 mg Hg/kg (SNTs grense for omsetning), og det ble funnet individer av denne størrelsen med konsentrasjoner opp mot 2,5 mg Hg/kg. -

Rapport 893/03 Utførende Institusjon NIVA
Oppdragsgivere Statens forurensningstilsyn Rapport 893/03 Utførende institusjon NIVA Kvikksølv i ferskvannsfisk fra Sør- Norge i 1998 – 2002, nivåer og tidsmessig utvikling TA-2000/2003 Norsk institutt for vannforskning Norsk institutt for vannforskning RAPPORT Hovedkontor Sørlandsavdelingen Østlandsavdelingen Vestlandsavdelingen Akvaplan-niva Postboks 173, Kjelsås Televeien 3 Sandvikaveien 41 Nordnesboder 5 0411 Oslo 4979 Grimstad 2312 Otestad 5008 Bergen 9296 Tromsø Telefon (47) 22 18 51 00 Telefon (47) 37 29 50 55 Telefon (47) 67 57 64 00 Telefon (47) 55 30 22 50 Telefon (47) 77 75 03 00 Telefax (47) 22 18 52 00 Telefax (47) 37 04 45 13 Telefax (47) 62 57 66 53 Telefax (47) 55 30 22 51 Telefax (47) 77 75 03 01 Internet: www.niva.no Tittel Løpenr. (for bestilling) Dato Kvikksølv i ferskvannsfisk fra Sør-Norge i 1998-2002, nivåer og 4813-2004 februar 2004 tidsmessig utvikling Prosjektnr. Undernr. Sider Pris O-21215 57 Forfattere Fagområde Distribusjon Eirik Fjeld og Sigurd Rognerud Miljøgifter Fri Geografisk område Trykket Norge NIVA Oppdragsgiver(e) Oppdragsreferanse Statens forurensningstilsyn (SFT) Per Erik Iversen Mattilsynet Are Sletta Sammendrag Rapporten sammenstiller data på kvikksølvkonsentrasjonene i ørret, abbor og gjedde fra 51 innsjøer i Sør-Norge, inn- fanget i periden 1998-2002. Typiske nivåer hos ørret av standardisert lengde (25 cm, 160 g) var 0,07– 0,14 mg Hg/kg. Hos abbor (21 cm, 100 g) var typiske konsentrasjoner i intervallet 0,17–0,48 mg/kg. Hos gjedde (53 cm, 1 kg) var de 0,41-0,81 mg/kg. Konsentrasjonene viste en tydelig tendens til å øke med fiskens lengde. -
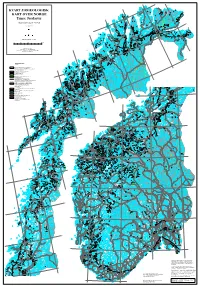
Jordarter V E U N O T N a Leirpollen
30°E 71°N 28°E Austhavet Berlevåg Bearalváhki 26°E Mehamn Nordkinnhalvøya KVARTÆRGEOLOGISK Båtsfjord Vardø D T a e n a Kjøllefjord a n f u j o v r u d o e Oksevatnet t n n KART OVER NORGE a Store L a Buevatnet k Geatnjajávri L s Varangerhalvøya á e Várnjárga f g j e o 24°E Honningsvåg r s d Tema: Jordarter v e u n o t n a Leirpollen Deanodat Vestertana Quaternary map of Norway Havøysund 70°N en rd 3. opplag 2013 fjo r D e a T g tn e n o a ra u a a v n V at t j j a n r á u V Porsanger- Vadsø Vestre Kjæsvatnet Jakobselv halvøya o n Keaisajávri Geassájávri o Store 71°N u Bordejávrrit v Måsvatn n n i e g Havvannet d n r evsbotn R a o j s f r r Kjø- o Bugøy- e fjorden g P fjorden 22°E n a Garsjøen Suolo- s r Kirkenes jávri o Mohkkejávri P Sandøy- Hammerfest Hesseng fjorden Rypefjord t Bjørnevatn e d n Målestokk (Scale) 1:1 mill. u Repparfjorden s y ø r ø S 0 25 50 100 Km Sørøya Sør-Varanger Sállan Skáiddejávri Store Porsanger Sametti Hasvik Leaktojávri Kartet inngår også i B áhèeveai- NASJONALATLAS FOR NORGE 20°E Leavdnja johka u a Lopphavet -

Stedsnavn I Nord-Østerdal
Stedsnavn i Nord-Østerdal JON OLAV RYEN Stedsnavn i Nord-Østerdal med Rondane, Rørosfjella og Femundstraktene KOLOFON FORLAG © Jon Olav Ryen Stedsnavn i Nord-Østerdal med Rondane, Rørosfjella og Femundstraktene Kolofon Forlag AS 2015 Forside- og baksidefoto: Rondane, med Midtronden og Digerronden (foto: Geir Olav Slåen). Prosjektet produseres på oppdrag fra forfatter/oppdragsgiver/utgiver Jon Olav Ryen Alle rettigheter/ansvar for prosjektets innhold tillegges Jon Olav Ryen Henvendelser utover bestilling av produktet bes rettet til Jon Olav Ryen ISBN 978-82-300-1287-1 Produksjon: Kolofon Forlag AS, 2015 Boken kan kjøpes i bokhandelen eller www.kolofon.com Det må ikke kopieres fra denne boken i strid med Åndsverkloven eller avtaler om kopiering inngått med KOPINOR, interesseorgan for rettighetshavere til åndsverk. Kopiering i strid med lov eller avtale medfører erstatningsansvar og inndraging og kan straffes med bøter eller fengsel. Innhold Forord ..................................................... 7 Innledning ............................................. 8 Bakgrunn, formål og metode ................. 8 Utvalg av stedsnavn ............................... 10 Skrivemåte og oppsett ............................ 12 Henvisninger, bruk av kilder ................ 13 Lydskrift .................................................. 14 Forkortelser, forklaringer ...................... 15 Stedsnavn .............................................. 17 Sørsamiske terrengord ....................... 483 Kilder .................................................... -

Samordning Av Lokaliteter Og Framtidige Utfordringer
Statlig program for forurensingsovervåking Nasjonale programmer for innsjøovervåking Samordning av lokaliteter og framtidige utfordringer TA-1949/2003 ISBN 82-577-4320-8 Referer til denne rapporten som: SFT, 2003. Nasjonale programmer for innsjøovervåking - Samordning av lokaliteter og framtidige utfordringer. (TA-1949/2003) Oppdragsgivere: Statens forurensningstilsyn Postboks 8100 Dep. 0032 Oslo Utførende institusjoner: Norsk institutt for naturforskning Tungasletta 2 7485 Trondheim Norsk institutt for vannforskning Postboks 173 Kjelsås 0411 Oslo Akvaplan-niva AS 9296 Tromsø Forord SFT har i e-mail 24.06.2002 anmodet NIVA om å koordinere et prosjektforslag i samarbeid med Akvaplan-niva og NINA angående fremtidig overvåking av innsjøer og koordinering av ulike overvåkingsprogrammer. Det er et ønske fra SFT om å samordne utvalget av innsjølokaliteter i seks nasjonale overvåkingsprogrammer og se dette i lys av framtidige planer for overvåking og i forbindelse med implementering av Vanndirektivet. Målsetningen er å skaffe en bedre oversikt over de aktivitetene som har foregått til nå, for om mulig å redusere kostnader ved fremtidig feltarbeid og innsamling. Videre ønsker SFT å dra nytte av informasjonen fra forskjellige overvåkingsprogrammer for å få større kunnskap om tilstanden i hver enkelt innsjø. Denne oversikten skal også være utgangspunkt for utvalg av lokaliteter til framtidig overvåking. Denne rapporten gir en oversikt over lokaliteter og aktiviteter i ca. 1000 lokaliteter i 6 nasjonale overvåkingsprogrammer og diskuterer mulighetene -
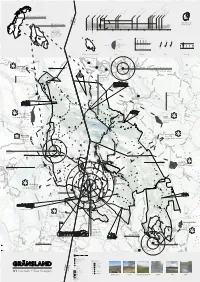
01 Context + Site Analysis
ing techniques d ish side orks orks ing by the Sami RIEN d os Copper W eindeer her os Copper W ing to come ines due to övelsjön opens oject e start ee l icated r ion Pr ease in farming on the Swed Rising tr 2009-2011 Gränslandet End of the last Ice Age Domest Mountain farms 1644 Start of Rør Falcon CatchingCarl Linneaus visits Gränslandet Incr Tourists ar 1926 DNT1937 Svukuriset STF GrChange opens in Sami lifestyle:1977 modern Closing housing of Røimte rand changeher cl Informat 01 Norway & Sweden 835 500 km2 Mountain farms Cattle herding Rogen Naturreservat is Logging and log driving Lumberjack cabins NORWAY home to Swedens only 02 Gränslandet SWEDEN tribe of wild muskox. 2 Ramundberget44 km radius 2000 km “circumference right” 9 areas of protected AURSUNDEN nature situated on the ago 1600 1700 1800 1900 2000 southern tip of the years Scandinavian Mountain 000 8000 years ago range, crossing over the 10 border of Norway and Brekken Messlingen Sweden. Røros 29.6 °C 30.1 °C 28.7 °C 29.3 °C SPRING mid April - mid June Tännäs Bruksvallarna Yearly average 1.6 °C 1.36 m ÖVER-SÄRVSJÖN Hitterdal WINTER SUMMER 1.0 m November - mid April mid June - September 0.5 m 1. 2.8 m/s 2. 8 m/s 3. 38.1 m/s 0.2 m -28.8 °C -31.4 °C SW 197° Fjällnäs -38 °C -37.1 °C l Drevsjø AUTUMN l os r Idre Fjäll September - November evsjø Rø ännäs r e Fjä r T D Id Weather stations for data collection Seasons Temperature Wind Snow depth Langtjønna FERAGEN Tänndalen LANDSKAPSVERNOMRÅDE Storvigeln Storvigeln since 2003 1561 MASL Fjällnäs, Tänndalen, Funäsdalen, Tännäs Highest mountain top Funäsdalen protected area OLD SAMI HUT Villages north of Gränslandet. -

Metodeutvikling: Overvåking Av Fisk I Store Innsjøer (FIST) I 2016
1573 Metodeutvikling: overvåking av fisk i store innsjøer (FIST) i 2016 Karl Øystein Gjelland, Odd Terje Sandlund, Oddgeir Andersen, Gunnbjørn Bremset, Knut Andreas E. Bækkelie, Jan Grimsrud Davidsen, Antti Eloranta, Oskar Pettersen, Lars Rønning, Atle Rustadbakken, Laila Saksgård, Randi Saksgård & Aslak Darre Sjursen NINAs publikasjoner NINA Rapport Dette er en elektronisk serie fra 2005 som erstatter de tidligere seriene NINA Fagrapport, NINA Oppdragsmelding og NINA Project Report. Normalt er dette NINAs rapportering til oppdragsgiver etter gjennomført forsknings-, overvåkings- eller utredningsarbeid. I tillegg vil serien favne mye av instituttets øvrige rapportering, for eksempel fra seminarer og konferanser, resultater av eget forsk- nings- og utredningsarbeid og litteraturstudier. NINA Rapport kan også utgis på annet språk når det er hensiktsmessig. NINA Kortrapport Dette er en enklere og ofte kortere rapportform til oppdragsgiver, gjerne for prosjekt med mindre arbeidsomfang enn det som ligger til grunn for NINA Rapport. Det er ikke krav om sammendrag på engelsk. Rapportserien kan også benyttes til framdriftsrapporter eller foreløpige meldinger til opp- dragsgiver. NINA Temahefte Som navnet angir behandler temaheftene spesielle emner. Heftene utarbeides etter behov og se- rien favner svært vidt; fra systematiske bestemmelsesnøkler til informasjon om viktige problemstil- linger i samfunnet. NINA Temahefte gis vanligvis en populærvitenskapelig form med mer vekt på illustrasjoner enn NINA Rapport. NINA Fakta Faktaarkene har som mål å gjøre NINAs forskningsresultater raskt og enkelt tilgjengelig for et større publikum. De sendes til presse, ideelle organisasjoner, naturforvaltningen på ulike nivå, politikere og andre spesielt interesserte. Faktaarkene gir en kort framstilling av noen av våre viktigste forsk- ningstema. Annen publisering I tillegg til rapporteringen i NINAs egne serier publiserer instituttets ansatte en stor del av sine viten- skapelige resultater i internasjonale journaler, populærfaglige bøker og tidsskrifter. -

Join the Worldwide HI Community Bli En Del Av HI-Familien!
About us Om oss Hostelling International Norway Hostelling International Norge • Non-profit membership organisation. • Ideell medlemsorganisasjon. • Proud history since 1932 and a relevant philosophy. • En stolt vandrerhjemshistorie siden 1932. Hostels in • Hostelling International is one of the world’s largest • Hostelling International er en av verdens største Norway 2018 youth membership organisations medlemsorganisasjoner for unge ay - 3.4 million members - 3,4 millioner medlemmer BecomeNorw a member of - Choice of 3,900 youth hostels worldwide, all of which - Et nettverk av 3,900 hosteller over hele verden, som meet internationally assured quality standards. BecomeHI Norwaya member of alle møter internasjonale standarder. • Much more than just a place to stay – we are working - Mye mer enn bare en seng – vi jobber for bedre Join us, getHI discounts Norway and support our towards better understanding between people by forståelse mellom mennesker gjennom å se nye steder, Becomenon-profit organisationa member that brings of discovering new places, new cultures and making oppleve nye kulturer og få venner for livet. Join us, getyoungHI discounts peopleNorway together! and support our lifelong friendships. non-profit organisation that brings • Vårt formål er å skape sosiale møteplasser. Dette gjør www.hihostels.no • Our mission is to create social meeting places by Join us, get discounts and support our vi ved å tilby rimelig overnatting for enkeltreisende, young people together! providing affordable accommodation for individuals, non-profit organisation that brings familier, grupper og organisasjoner. www.hihostels.no families, groups and organisations. young people together! • Hos oss er alle velkomne, uansett kulturell bakgrunn, • Everyone is welcome, regardless of cultural back- www.hihostels.no hudfarge eller religiøs tilhørighet. -
Femundsmarka
FEMUNDSMARKA With a canoe and a fishing rod to Norway’s morning land 2° 3° Femundsmarka National Park Femundsmarka National Park The gift from the ice With twisted pines and secluded lakes scattered amongst a sea of boulders, the Femundsmarka National Park beckons you to experience it. Ten thousand years ago, the glaciers retreated leaving behind a desolate, primeval-like, ice-age landscape – a landscape that has scarcely changed since. Here you can paddle a canoe for days along the many rivers and lakes. You can take a fishing rod with you to catch the large trout that can be found here, and if you keep your eyes open to the beautiful landscape, you will be rewarded with a memorable trip. The area has been used by the Sámi people for centuries, but retains its unspoilt character. Together with adjoining protected areas in Sweden, the Femundsmarka National Park forms part of one of the largest continuous, undisturbed wilderness areas in southern Scandinavia. It is also an area that provides a habitat for a number of rare and vulnerable animal and bird species. Autumnal colouring in Femundsmarka (BL) 4° 5° Femundsmarka National Park Femundsmarka National Park Canoeing (BL) Evening light on upland downy birches (BL) ENJOY THE SCENERY About half the visitors to the Femundsmarka National Take a trip! Park go canoeing or fishing. The area is also spectacular For hunting, berry-picking, skiing or hiking, Femunds- for hiking - whether you want to go hunting or berry- marka offers plenty of options. During the summer picking, or just enjoy the scenery. -

Nasjonalt Referansesystem for Landskap Beskrivelse Av Norges 45 Landskapsregioner Oskar Puschmann
Nasjonalt referansesystem for landskap Beskrivelse av Norges 45 landskapsregioner Oskar Puschmann NIJOS rapport 10/2005 Nasjonalt referansesystem for landskap - beskrivelse av Norges 45 landskapsregioner av Oskar Puschmann Forsidefoto: Oskar Puschmann Heimlandet på Røst, Røst kommune, Nordland. Landskapsregion 30 Nordlandsverran. NIJOS rapport 10/2005 Tittel: Nasjonalt referansesystem for landskap. NIJOS nummer: Beskrivelse av Norges 45 landskapsregioner 10 /2005 Forfatter(e): ISBN nummer: Oskar Puschmann 82-7464-355-0 Oppdragsgiver: Dato: 13. des. 2005 Landbruks- og matdepartementet Prosjekt/Program: Nasjonalt referansesystem for landskap (RSL) Relatert informasjon/Andre publikasjoner i utvalg fra prosjektet: - Puschmann, O., Reid, S.J., Fjellstad, W., Hofsten, J. & Dramstad, W. 2004. Tilstandsbeskrivelse av norske jordbruksregioner ved bruk av statistikk. NIJOS-rapport 17/04. - Puschmann, O. 1998. Nasjonalt referansesystem for landskap. Bruk av ulike kilder som grunnlag for beskrivelse av underregioner. NIJOS-rapport 12/98. - Kamfjord, G., Lykkja, H. & Puschmann, O. 1997. Landskapet og reiselivsproduktet. NIJOS-rapport 04/97. - Elgersma, A. 1996. ”Norske landskapsregioner. Kart M 1: 2 mill.” Norsk institutt for jord og skogkartlegging.” Utdrag: Siden 1989 har NIJOS arbeidet med utviklingen av et nasjonalt referansesystem for landskap. Dette er et hierarkisk system med inndeling av landet i 45 landskapsregioner og 444 underregioner. Med utgangspunkt i underregionene kan det videre avgrenses i landskapsområder på lokalt nivå. I denne rapporten gis en innføring i metoden som ligger til grunn. Videre inneholder rapporten en beskrivelse, samt visualisering med kart og bilder, av hver av de 45 landskapsregionene. Beskrivelsene skildrer seks ulike landskapskomponenter; landskapets hovedform, landskapets småformer, vann og vassdrag, vegetasjon, jordbruksmark og bebyggelse og tekniske anlegg. I tillegg følger en beskrivelse av regionens samlede landskapskarakter, og her er det også belyst enkelte særegne regionale kvaliteter, problemområder eller utviklingstrender. -

Fiskeguide Roasten 7 12 Stortjønn 5 4
Røros 3 ENGERDAL Røvola Nedre Fiskeguide Roasten 7 12 Stortjønn 5 4 2 Øvre 8 1 Femund Ø, R, H, S, A, G 65 Femundshytta Røvoltjønnan Roasten Storfisk- 2 Røvoltjerna Ø, R, A 6 tjønnan Buhøgda Litle 783 3 Nedre Røa Ø, H, G 4 Vonsjøen 11 Haugen gård 4 Skogtjern Ø, R Litlevonsjø- Det gjøres oppmerksom på at det er en Grøtåa vola Korstjern Ø, R mengde tjern, åer og bekker med fine fiskebe- stander som ikke er omtalt eller kommer fram i 9 5 Roastsjøene Ø, R, H, A, G fiskekartet. Ugland ITs turkartserie i målestokk Grøtådalsætra 10 788 6 Grøtåa Ø 1:50 000, kartblad Femund nord, Femund sør Kuvolsætra Vonsjøen 7 Øvre Røa Ø, R, H, G, A og Engerdal dekker hele kommunen og kan 1416 Grøtådalen 8 Storfisktjern Ø, R, H, G, A anbefales. Flata Stor-Svuku 9 Kratla Ø, R, A Ø = Ørret Svukuris- 10 Store Vonsjøen Ø, R, A, G R = Røye Jonasvollen Rønsjø- Svukuriset Litl-Svuku 11 Lille Vonsjøen R, A H = Harr floan ruten 12 Rogen Ø, R, H, A, G S = Sik Revlinga 13 Rønsjøen Ø, R A = Abbor 13 Revling- 888 Rønsjøen G = Gjedde 14 81 14 Revlingsjøen Ø, R sjøan Røa M = Mort 1 Sandtjønn- 15 Lille Grøvelsjøen Ø 16 Elgåsjøen Ø, R, A vola 17 Volsjøen Ø, R Korneset Øyan Os Sylen 18 Hansbekktjern Ø, R Røsten Elgå Ryvang 82 19 Djupsjøen Ø, R, A Egga Grøvelsjøen Sylens private område Litlå- Kvernvika Elgåhogna 20 Båthussjøen Ø, A 1460 Tolga vollen Solli 21 Storbekktjerna Ø Litlgrøvelsjø- Sals- 22 Flåtesjøen Ø Gjotsætra Johnsgård Sjøli Dalset 15 floan fjellet 23 Skogtjern Ø,R 52 Tjønnodd- Buvika 16 Digerhogna 24 Storsjøen Ø,R,A Langsjøen vola 17 25 Hammersjøen Ø,R,A 18 Volsjøen