Sox10 Has a Broad Expression Pattern in Gliomas and Enhances Platelet-Derived Growth Factor-B–Induced Gliomagenesis
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Neurofibromatosis Type 2 (NF2)
International Journal of Molecular Sciences Review Neurofibromatosis Type 2 (NF2) and the Implications for Vestibular Schwannoma and Meningioma Pathogenesis Suha Bachir 1,† , Sanjit Shah 2,† , Scott Shapiro 3,†, Abigail Koehler 4, Abdelkader Mahammedi 5 , Ravi N. Samy 3, Mario Zuccarello 2, Elizabeth Schorry 1 and Soma Sengupta 4,* 1 Department of Genetics, Cincinnati Children’s Hospital, Cincinnati, OH 45229, USA; [email protected] (S.B.); [email protected] (E.S.) 2 Department of Neurosurgery, University of Cincinnati, Cincinnati, OH 45267, USA; [email protected] (S.S.); [email protected] (M.Z.) 3 Department of Otolaryngology, University of Cincinnati, Cincinnati, OH 45267, USA; [email protected] (S.S.); [email protected] (R.N.S.) 4 Department of Neurology, University of Cincinnati, Cincinnati, OH 45267, USA; [email protected] 5 Department of Radiology, University of Cincinnati, Cincinnati, OH 45267, USA; [email protected] * Correspondence: [email protected] † These authors contributed equally. Abstract: Patients diagnosed with neurofibromatosis type 2 (NF2) are extremely likely to develop meningiomas, in addition to vestibular schwannomas. Meningiomas are a common primary brain tumor; many NF2 patients suffer from multiple meningiomas. In NF2, patients have mutations in the NF2 gene, specifically with loss of function in a tumor-suppressor protein that has a number of synonymous names, including: Merlin, Neurofibromin 2, and schwannomin. Merlin is a 70 kDa protein that has 10 different isoforms. The Hippo Tumor Suppressor pathway is regulated upstream by Merlin. This pathway is critical in regulating cell proliferation and apoptosis, characteristics that are important for tumor progression. -

Central Nervous System Cancers Panel Members Can Be Found on Page 1151
1114 NCCN David Tran, MD, PhD; Nam Tran, MD, PhD; Frank D. Vrionis, MD, MPH, PhD; Patrick Y. Wen, MD; Central Nervous Nicole McMillian, MS; and Maria Ho, PhD System Cancers Overview In 2013, an estimated 23,130 people in the United Clinical Practice Guidelines in Oncology States will be diagnosed with primary malignant brain Louis Burt Nabors, MD; Mario Ammirati, MD, MBA; and other central nervous system (CNS) neoplasms.1 Philip J. Bierman, MD; Henry Brem, MD; Nicholas Butowski, MD; These tumors will be responsible for approximately Marc C. Chamberlain, MD; Lisa M. DeAngelis, MD; 14,080 deaths. The incidence of primary brain tumors Robert A. Fenstermaker, MD; Allan Friedman, MD; Mark R. Gilbert, MD; Deneen Hesser, MSHSA, RN, OCN; has been increasing over the past 30 years, especially in Matthias Holdhoff, MD, PhD; Larry Junck, MD; elderly persons.2 Metastatic disease to the CNS occurs Ronald Lawson, MD; Jay S. Loeffler, MD; Moshe H. Maor, MD; much more frequently, with an estimated incidence ap- Paul L. Moots, MD; Tara Morrison, MD; proximately 10 times that of primary brain tumors. An Maciej M. Mrugala, MD, PhD, MPH; Herbert B. Newton, MD; Jana Portnow, MD; Jeffrey J. Raizer, MD; Lawrence Recht, MD; estimated 20% to 40% of patients with systemic cancer Dennis C. Shrieve, MD, PhD; Allen K. Sills Jr, MD; will develop brain metastases.3 Abstract Please Note Primary and metastatic tumors of the central nervous system are The NCCN Clinical Practice Guidelines in Oncology a heterogeneous group of neoplasms with varied outcomes and (NCCN Guidelines®) are a statement of consensus of the management strategies. -

Malignant Glioma Arising at the Site of an Excised Cerebellar Hemangioblastoma After Irradiation in a Von Hippel-Lindau Disease Patient
DOI 10.3349/ymj.2009.50.4.576 Case Report pISSN: 0513-5796, eISSN: 1976-2437 Yonsei Med J 50(4): 576-581, 2009 Malignant Glioma Arising at the Site of an Excised Cerebellar Hemangioblastoma after Irradiation in a von Hippel-Lindau Disease Patient Na-Hye Myong1 and Bong-Jin Park2 1Department of Pathology, Dankook University College of Medicine, Cheonan; 2Department of Neurosurgery, Kyunghee University Hospital, Seoul, Korea. We describe herein a malignant glioma arising at the site of the resected hemangioblastoma after irradiation in a patient with von Hippel-Lindau disease (VHL). The patient was a 25 year-old male with multiple heman- gioblastomas at the cerebellum and spinal cord, multiple pancreatic cysts and a renal cell carcinoma; he was diagnosed as having VHL disease. The largest hemangioblastoma at the right cerebellar hemisphere was completely removed, and he received high-dose irradiation postoperatively. The tumor recurred at the same site 7 years later, which was a malignant glioma with no evidence of hemangioblastoma. The malignant glioma showed molecular genetic profiles of radiation-induced tumors because of its diffuse p53 immunostaining and the loss of p16 immunoreactivity. The genetic study to find the loss of heterozygosity (LOH) of VHL gene revealed that only the cerebellar hemangioblastoma showed allelic losses for the gene. To the best of our knowledge, this report is the first to show a malignant glioma that developed in a patient with VHL disease after radiation therapy at the site of an excised hemangioblastoma. This report also suggests that radiation therapy should be performed very carefully in VHL patients with hemangioblastomas. -

Integrated and Functional Genomic Approaches to Elucidate Differential Genetic Dependencies in Melanoma
Integrated and Functional Genomic Approaches to Elucidate Differential Genetic Dependencies in Melanoma The Harvard community has made this article openly available. Please share how this access benefits you. Your story matters Citation Wong, Terence. 2018. Integrated and Functional Genomic Approaches to Elucidate Differential Genetic Dependencies in Melanoma. Doctoral dissertation, Harvard University, Graduate School of Arts & Sciences. Citable link http://nrs.harvard.edu/urn-3:HUL.InstRepos:42014990 Terms of Use This article was downloaded from Harvard University’s DASH repository, and is made available under the terms and conditions applicable to Other Posted Material, as set forth at http:// nrs.harvard.edu/urn-3:HUL.InstRepos:dash.current.terms-of- use#LAA Integrated and Functional Genomic Approaches to Elucidate Differential Genetic Dependencies in Melanoma A dissertation presented by Terence Cheng Wong to The Division of Medical Sciences in partial fulfillment of the requirements for the degree of Doctor of Philosophy in the subject of Biological and Biomedical Sciences Harvard University Cambridge, Massachusetts November 2017 © 2017 Terence Cheng Wong All rights reserved. Dissertation Advisor: Levi Garraway Terence Cheng Wong Integrated and Functional Genomic Approaches to Elucidate Differential Genetic Dependencies in Melanoma ABSTRACT Genomic characterization of human cancers over the past decade has generated comprehensive catalogues of genetic alterations in cancer genomes. Many of these genetic events result in molecular or cellular changes that drive cancer cell phenotypes. In melanoma, a majority of tumors harbor mutations in the BRAF gene, leading to activation of the MAPK pathway and tumor initiation. The development and use of drugs that target the mutant BRAF protein and the MAPK pathway have produced significant clinical benefit in melanoma patients. -

Charts Chart 1: Benign and Borderline Intracranial and CNS Tumors Chart
Charts Chart 1: Benign and Borderline Intracranial and CNS Tumors Chart Glial Tumor Neuronal and Neuronal‐ Ependymomas glial Neoplasms Subependymoma Subependymal Giant (9383/1) Cell Astrocytoma(9384/1) Myyppxopapillar y Desmoplastic Infantile Ependymoma Astrocytoma (9412/1) (9394/1) Chart 1: Benign and Borderline Intracranial and CNS Tumors Chart Glial Tumor Neuronal and Neuronal‐ Ependymomas glial Neoplasms Subependymoma Subependymal Giant (9383/1) Cell Astrocytoma(9384/1) Myyppxopapillar y Desmoplastic Infantile Ependymoma Astrocytoma (9412/1) (9394/1) Use this chart to code histology. The tree is arranged Chart Instructions: Neuroepithelial in descending order. Each branch is a histology group, starting at the top (9503) with the least specific terms and descending into more specific terms. Ependymal Embryonal Pineal Choro id plexus Neuronal and mixed Neuroblastic Glial Oligodendroglial tumors tumors tumors tumors neuronal-glial tumors tumors tumors tumors Pineoblastoma Ependymoma, Choroid plexus Olfactory neuroblastoma Oligodendroglioma NOS (9391) (9362) carcinoma Ganglioglioma, anaplastic (9522) NOS (9450) Oligodendroglioma (9390) (9505 Olfactory neurocytoma Ganglioglioma, malignant (()9521) anaplastic (()9451) Anasplastic ependymoma (9505) Olfactory neuroepithlioma Oligodendroblastoma (9392) (9523) (9460) Papillary ependymoma (9393) Glioma, NOS (9380) Supratentorial primitive Atypical EdEpendymo bltblastoma MdllMedulloep ithliithelioma Medulloblastoma neuroectodermal tumor tetratoid/rhabdoid (9392) (9501) (9470) (PNET) (9473) tumor -

Central Nervous System Tumors General ~1% of Tumors in Adults, but ~25% of Malignancies in Children (Only 2Nd to Leukemia)
Last updated: 3/4/2021 Prepared by Kurt Schaberg Central Nervous System Tumors General ~1% of tumors in adults, but ~25% of malignancies in children (only 2nd to leukemia). Significant increase in incidence in primary brain tumors in elderly. Metastases to the brain far outnumber primary CNS tumors→ multiple cerebral tumors. One can develop a very good DDX by just location, age, and imaging. Differential Diagnosis by clinical information: Location Pediatric/Young Adult Older Adult Cerebral/ Ganglioglioma, DNET, PXA, Glioblastoma Multiforme (GBM) Supratentorial Ependymoma, AT/RT Infiltrating Astrocytoma (grades II-III), CNS Embryonal Neoplasms Oligodendroglioma, Metastases, Lymphoma, Infection Cerebellar/ PA, Medulloblastoma, Ependymoma, Metastases, Hemangioblastoma, Infratentorial/ Choroid plexus papilloma, AT/RT Choroid plexus papilloma, Subependymoma Fourth ventricle Brainstem PA, DMG Astrocytoma, Glioblastoma, DMG, Metastases Spinal cord Ependymoma, PA, DMG, MPE, Drop Ependymoma, Astrocytoma, DMG, MPE (filum), (intramedullary) metastases Paraganglioma (filum), Spinal cord Meningioma, Schwannoma, Schwannoma, Meningioma, (extramedullary) Metastases, Melanocytoma/melanoma Melanocytoma/melanoma, MPNST Spinal cord Bone tumor, Meningioma, Abscess, Herniated disk, Lymphoma, Abscess, (extradural) Vascular malformation, Metastases, Extra-axial/Dural/ Leukemia/lymphoma, Ewing Sarcoma, Meningioma, SFT, Metastases, Lymphoma, Leptomeningeal Rhabdomyosarcoma, Disseminated medulloblastoma, DLGNT, Sellar/infundibular Pituitary adenoma, Pituitary adenoma, -

A Rare Disease in Pediatric Patients
Cartas ao Editor Jornal de Pediatria - Vol. 85, Nº 3, 2009 277 Resposta do autor Gliomatose leptomeníngea primária difusa: uma doença rara em pacientes pediátricos Prezado Editor, Recebemos com prazer os comentários acima. Os autores esclarecem que, em momento algum, focaram suas preo- Prezado Editor, cupações em critérios para definir síndrome metabólica, até mesmo porque se ela existe1, não existem critérios de consenso O recente relato de caso de Val Filho & Avelar1 sobre um para o diagnóstico em adultos2,3 tampouco em adolescentes. raro tumor cerebral pediátrico é interessante e merece grande Preocupamo-nos sim, em mostrar a presença de fatores de consideração no contexto da literatura publicada sobre esse risco em uma amostra selecionada ao acaso, que eles existem assunto, já que representa um evento muito incomum. Em em proporções tais que podem até ser agrupados, e, por um nome da precisão científica, devemos, portanto, fazer uma dos muitos critérios sugeridos na literatura, com valores de importante correção no que se refere ao diagnóstico. referências pediátricos, dar origem ao que se denomina sín- Os autores fizeram uma revisão breve e adequada sobre a drome metabólica. Não nos propusemos, portanto, a estudar gliomatose cerebral; uma doença rara, classificada pela edição associações entre fatores de risco. de 2007 da Organização Mundial da Saúde (OMS)2 como uma Os autores acreditam que discutir critérios para o que não lesão de grau III com prognóstico reservado. A gliomatose 2,3 existe consenso é desfocar a atenção da gravidade -

6 Magnetic Resonance Imaging
Chapter 6 / MRI 105 6 Magnetic Resonance Imaging Paul M. Ruggieri Summary Magnetic resonance (MR) is clearly the accepted imaging standard for the preliminary evalua- tion, peri-operative management, and routine longitudinal follow-up of patients with high-grade glio- mas (HGG). The purpose of this chapter is to review the imaging characteristics of HGG using conventional MR imaging techniques. Whereas the newer techniques of MR diffusion, perfusion, diffusion tensor imaging, and MR spectroscopy will be included as part of this discussion of the high grade neoplasms, the detailed concepts of such studies will be discussed elsewhere in this text. Key Words: MR imaging; anaplastic astrocytoma; glioblastoma multiforme; gliosarcoma; gliomatosis cerebri; oligodendroglioma. INTRODUCTION Magnetic resonance (MR) is clearly the accepted imaging standard for the preliminary evaluation, peri-operative management, and routine longitudinal follow-up of patients with high-grade gliomas (HGG). The purpose of this chapter is to review the imaging characteristics of HGG using conventional MR imaging techniques. Whereas the newer techniques of MR diffusion, perfusion, diffusion tensor imaging, and MR spectroscopy will be included as part of this discussion of the high grade neoplasms, the detailed concepts of such studies will be discussed elsewhere in this text. In general terms, high-grade glial neoplasms are conventionally thought of as infiltrative parenchymal masses that are hyperintense on FLAIR fluid-attenuated inversion recovery (FLAIR) and T2-weighted images, hypointense on unenhanced T1-weighted images, may or may not extend into the corpus callosum, are surrounded by extensive vasogenic edema, and prominently enhance following gadolinium administration. It must be pointed out that this description is clearly just a generalization as many high-grade neoplasms clearly do not follow these “rules” and some of these characteristics may even be seen in low-grade neoplasms. -

Olig1 and Sox10 Interact Synergistically to Drivemyelin Basic
The Journal of Neuroscience, December 26, 2007 • 27(52):14375–14382 • 14375 Cellular/Molecular Olig1 and Sox10 Interact Synergistically to Drive Myelin Basic Protein Transcription in Oligodendrocytes Huiliang Li,1 Yan Lu,2 Hazel K. Smith,1 and William D. Richardson1 1Wolfson Institute for Biomedical Research and Department of Biology, University College London, London WC1E 6BT, United Kingdom, and 2Medical Research Council, Clinical Sciences Centre, Imperial College London, London W12 0NN, United Kingdom The oligodendrocyte lineage genes (Olig1/2), encoding basic helix-loop-helix transcription factors, were first identified in screens for master regulators of oligodendrocyte development. OLIG1 is important for differentiation of oligodendrocyte precursors into myelin- forming oligodendrocytes during development and is thought to play a crucial role in remyelination during multiple sclerosis. However, itisstillunclearhowOLIG1interactswithitstranscriptionalcofactorsandDNAtargets.OLIG1wasreportedlyrestrictedtomammals,but we demonstrate here that zebrafish and other teleosts also possess an OLIG1 homolog. In zebrafish, as in mammals, Olig1 is expressed in the oligodendrocyte lineage. Olig1 associates physically with another myelin-associated transcription factor, Sox10, and the Olig1/Sox10 complex activates mbp (myelin basic protein) transcription via conserved DNA sequence motifs in the mbp promoter region. In contrast, Olig2 does not bind to Sox10 in zebrafish, although both OLIG1 and OLIG2 bind SOX10 in mouse. Key words: Olig1; Olig2; Sox10; Mbp; oligodendrocyte; myelin; zebrafish; mouse; evolution; development Introduction directly regulates Mbp transcription (Stolt et al., 2002), and over- Myelin, the multilayered glial sheath around axons, is one of the expression of SOX10 alone is sufficient to induce myelin gene defining features of jawed vertebrates (gnathostomes). It is expression in embryonic chick spinal cord (Liu et al., 2007). -
A Case of Intramedullary Spinal Cord Astrocytoma Associated with Neurofibromatosis Type 1
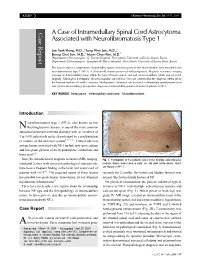
KISEP J Korean Neurosurg Soc 36 : 69-71, 2004 Case Report A Case of Intramedullary Spinal Cord Astrocytoma Associated with Neurofibromatosis Type 1 Jae Taek Hong, M.D.,1 Sang Won Lee, M.D.,1 Byung Chul Son, M.D.,1 Moon Chan Kim, M.D.2 Department of Neurosurgery,1 St. Vincent Hospital, The Catholic University of Korea, Suwon, Korea Department of Neurosurgery,2 Kangnam St. Mary's Hospital, The Catholic University of Korea, Seoul, Korea The authors report a symptomatic intramedullary spinal cord astrocytoma in the thoracolumbar area associated with neurofibromatosis type 1 (NF-1). A 38-year-old woman presented with paraparesis. Magnetic resonance imaging revealed an intramedullary lesion within the lower thoracic spinal cord and conus medullaris, which was removed surgically. Pathological investigation showed anaplastic astrocytoma. This case confirms that the diagnosis criteria set by the National Institute of Health Consensus Development Conference can be useful to differentiate ependymoma from astrocytoma when making a preoperative diagnosis of intramedullary spinal cord tumor in patients of NF-1. KEY WORDS : Astrocytoma·Intramedullary cord tumor·Neurofibromatosis. Introduction eurofibromatosis type 1 (NF-1), also known as von N Recklinghausen's disease, is one of the most common autosomal dominant inherited disorders with an incidence of 1 in 3,000 individuals and is characterized by a predisposition to tumors of the nervous system5,6,12,16). Central nervous system lesions associated with NF-1 include optic nerve glioma and low-grade gliomas of the hypothalamus, cerebellum and brain stem6,10). Since the introduction of magnetic resonance(MR) imaging, Fig. 1. Photograph of the patient's back shows multiple subcutaneous incidental lesions with uncertain pathological characteristic nodules (black arrow) and a cafe-au-lait spot (white arrow), which have been a frequent finding in the brain and spinal cord of are typical of NF-1. -

SUPPLEMENTARY MATERIAL Bone Morphogenetic Protein 4 Promotes
www.intjdevbiol.com doi: 10.1387/ijdb.160040mk SUPPLEMENTARY MATERIAL corresponding to: Bone morphogenetic protein 4 promotes craniofacial neural crest induction from human pluripotent stem cells SUMIYO MIMURA, MIKA SUGA, KAORI OKADA, MASAKI KINEHARA, HIROKI NIKAWA and MIHO K. FURUE* *Address correspondence to: Miho Kusuda Furue. Laboratory of Stem Cell Cultures, National Institutes of Biomedical Innovation, Health and Nutrition, 7-6-8, Saito-Asagi, Ibaraki, Osaka 567-0085, Japan. Tel: 81-72-641-9819. Fax: 81-72-641-9812. E-mail: [email protected] Full text for this paper is available at: http://dx.doi.org/10.1387/ijdb.160040mk TABLE S1 PRIMER LIST FOR QRT-PCR Gene forward reverse AP2α AATTTCTCAACCGACAACATT ATCTGTTTTGTAGCCAGGAGC CDX2 CTGGAGCTGGAGAAGGAGTTTC ATTTTAACCTGCCTCTCAGAGAGC DLX1 AGTTTGCAGTTGCAGGCTTT CCCTGCTTCATCAGCTTCTT FOXD3 CAGCGGTTCGGCGGGAGG TGAGTGAGAGGTTGTGGCGGATG GAPDH CAAAGTTGTCATGGATGACC CCATGGAGAAGGCTGGGG MSX1 GGATCAGACTTCGGAGAGTGAACT GCCTTCCCTTTAACCCTCACA NANOG TGAACCTCAGCTACAAACAG TGGTGGTAGGAAGAGTAAAG OCT4 GACAGGGGGAGGGGAGGAGCTAGG CTTCCCTCCAACCAGTTGCCCCAAA PAX3 TTGCAATGGCCTCTCAC AGGGGAGAGCGCGTAATC PAX6 GTCCATCTTTGCTTGGGAAA TAGCCAGGTTGCGAAGAACT p75 TCATCCCTGTCTATTGCTCCA TGTTCTGCTTGCAGCTGTTC SOX9 AATGGAGCAGCGAAATCAAC CAGAGAGATTTAGCACACTGATC SOX10 GACCAGTACCCGCACCTG CGCTTGTCACTTTCGTTCAG Suppl. Fig. S1. Comparison of the gene expression profiles of the ES cells and the cells induced by NC and NC-B condition. Scatter plots compares the normalized expression of every gene on the array (refer to Table S3). The central line -

Birth Defects Caused by Mutations in Human GLI3 and Mouse Gli3 Genescga
doi:10.1111/j.1741-4520.2009.00266.x Congenital Anomalies 2010; 50, 1–7 1 REVIEW ARTICLE Birth defects caused by mutations in human GLI3 and mouse Gli3 genescga_ 266 1..71..7 Ichiro Naruse1, Etsuko Ueta1, Yoshiki Sumino1, Masaya Ogawa1, and Satoshi Ishikiriyama2 1School of Health Science, Faculty of Medicine, Tottori University, Yonago, and 2Division of Clinical Genetics and Cytogenetics, Shizuoka Children’s Hospital, Shizuoka, Japan ABSTRACT GLI3 is the gene responsible for Greig cepha- type-A (PAP-A) and preaxial polydactyly type-IV (PPD-IV). A lopolysyndactyly syndrome (GCPS), Pallister–Hall syndrome mimic phenomenon is observed in mice. Investigation of human (PHS) and Postaxial polydactyly type-A (PAP-A). Genetic poly- GLI3 and Gli3 genes has progressed using the knowledge of dactyly mice such as Pdn/Pdn (Polydactyly Nagoya), XtH/XtH Cubitus interruptus (Ci) in Drosophila, a homologous gene of GLI3 (Extra toes) and XtJ/XtJ (Extra toes Jackson) are the mouse and Gli3 genes. These genes have been highly conserved in the homolog of GCPS, and Gli3tmlUrtt/Gli3tmlUrt is produced as the animal kingdom throughout evolution. Now, a lot of mutant mice of mouse homolog of PHS. In the present review, relationships Gli3 gene have been known and knockout mice have been pro- between mutation points of GLI3 and Gli3, and resulting phe- duced. It is expected that the knowledge obtained from mutant and notypes in humans and mice are described. It has been con- knockout mice will be extrapolated to the manifestation mecha- firmed that mutation in the upstream or within the zinc finger nisms in human diseases to understand the diseases caused by the domain of the GLI3 gene induces GCPS; that in the post-zinc mutations in GLI3 gene.