Please Scroll Down for Article
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Review and Status of Biological Control of Clidemia in Hawaici
REVIEW AND STATUS OF BIOLOGICAL CONTROL OF CLIDEMIA IN HAWAICI Larry M. Nakahara, Robert Me Burkhart, and George Ye Funasaki ABSTRACT Efforts to control clidemia (Clidemia hirta) in Hawai'i with phytophagous insects began in 1952. Several attempts have been made since then to introduce potential biological control agents to reduce the spread of this weed into forest areas, and other control measures have been tried by various groups and agencies. It was not until recently, however, through the enactment of significant legislation and subsequent funding, that explorations and studies were conducted specifically on clidemia insects in Trinidad, West Indies. Fourteen species of insects were evaluated, including Carposina bullata, Mompha bithalama, a midge or cecidomyiid, two Eurytoma species, Piesmopoda sp., and Compsolechia seductella, which feed on the flowers and fruits; Lius poseidon, Antiblemma acclinalis, Druentia sp., prob. inscita, Ategumia matutinalis (formerly Blepharomastix ebulealis but originally thought to be Sylepte matutinalis), Penestes n. sp., and a leaf beetle or chrysomelid, which feed on the leaves; and a long-horned cerambycid beetle, which bores into the stem. Studies indicated that several species were sufficiently host specific to warrant introduction into Hawai'i for the biological control of clidemia. INTRODUCTION In Hawai'i, clidemia or Koster's curse (Clidemia hirta) (Melastomataceae), a fast-growing, hea -seeding, tropical American shrub, has colonized forest clearings, trailsi7 es, and bum sites and intruded into the understories of forests that were formerly free of introduced, or alien, weeds (Wester and Wood 1977). The species has spread rapidly throughout the State and re resents a serious threat to our native forests. -

(Lepidoptera: Noctuidae) on Miconia Calvescens DC
Biocon trol Science and Technology. ,,Taylor&Francis 1yIor & F—r Group Vol. 18, Nos. 1/2, 2008, 183 192 Biology, herbivory, and host specificity of Antibleinma leucocyma (Lepidoptera: Noctuidae) on Miconia calvescens DC. (Melastomataceae) in Brazil F.R. BadenesPerez I and M.T. Johnsonh "Pacific Cooperative Studies Unit, Universit y of Hawaii at Manoci, Honolulu, HI, USA; "Institute of Pacific Islands Forestry, USDA Forest Service Pacific Southwest Research Station, Vlcano, H!, USA Received 17 October 2007; final version received 11 December 2007) Miconia calvescens DC. (Melastomataceae) is an invasive tree considered one of the greatest threats to natural ecosystems of Hawaii and other Pacific islands. The potential for using the defoliator Antihiemma leucoc ynia (Lepidoptera: Noctui- dae) as a biological control agent of M. calvescens was evaluated in the native habitat of the insect in Brazil. Impact assessment in the field showed that 37.2/u of leaves presented damage by A. leucocyma. and among damaged leaves, 1.5-51.9% of the leaf area (16.3% on average) was affected. Damage was found in young to fully expanded leaves (28-915 cm in area). Individual A. leucocyrna developing as third through sixth instars in the laboratory consumed an average of 1348 cm of M. calvescens leaf material in about 11 days. Populations of A. leuco 1,117a in the field were heavily impacted (up to 83% parasitism) by a braconid parasitoid.' Observations in the field and single- and two-host choice tests in the laboratory indicated that A. leucocyrna has a narrow host range restricted to M. calvescens. Usefulness of this herbivore for biological control in Hawaii appears low, however, because of its probable susceptibility to generalist parasitoids. -

Two Species of the Mompha Trithalama-Complex As Possible Biological Control of Koster's Curse and Velvet Tree
Two species of the Mompha trithalama-complex as possible biological control of Koster’s curse and Velvet tree Sjaak Koster, Manuel Alfaro Alpízar, Francisco R. Badenes-Pérez & Kenji Nishida Mompha spec. A Mompha trithalama Mompha spec. B Mompha trithalama Mompha spec. A Mompha spec. B Mompha trithalama Mompha spec. A Clidemia hirta (L.) D.Don (Melastomataceae) Koster’s curse or soap bush Photo: www.cybertruffle.org.uk/vinales/esp/clidemia_hirta Clidemia hirta (L.) D.Don (Melastomataceae) Koster’s curse or soap bush Photo courtesy Konrad Englberger, © Secretariat of the Pacific Community Geographical distribution of Clidemia hirta in areas where it is native (circles) and introduced (squaires) (DeWalt, 2003) Damage to the native flora Photo courtesy Konrad Englberger, © Secretariat of the Pacific Community Damage to the native flora Clidemia hirta on Babeldaob, Palau Photo by Jim Space, PIER Attempts for biological control so far The fungus: Colletotrichum gloesporioides Penz. (Phyllachorales: Phyllachoraceae) The thrips: Liothrips urichi Karny (Thysanoptera: Phlaeothripidae) The beetle: Lius poseidon Napp (Coleoptera: Buprestidae) The moths: Antiblemma acclinalis Hübner (Noctuidae) Carposina bullata Meyrick (Carposinidae) Mompha trithalama Meyrick (Momphidae) Miconia calvescens Photo’s by The Nature Conservancy Archive, The Nature Conservancy Photo’s Wikipedia and http://pbin.nbii.gov/reportapest/maui/pestlist/miccal.htm Mark that the larva made when it got into the berry Photo Manuel Alfaro Alpízar Larva inside the berry Photo Manuel Alfaro Alpízar Fruits of Miconia calvescens with the larva of Mompha spec. A Photo Manuel Alfaro Alpízar Different instars of larvae of Mompha spec. A Photo Manuel Alfaro Alpízar Construction of cocoon Photo Manuel Alfaro Alpízar Thank you for your attention Abstract of my presentation held on the XV European Congress of Lepidopterology at Berlin from 8 – 12 september 2007. -

A List of Cuban Lepidoptera (Arthropoda: Insecta)
TERMS OF USE This pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website is prohibited. Zootaxa 3384: 1–59 (2012) ISSN 1175-5326 (print edition) www.mapress.com/zootaxa/ Article ZOOTAXA Copyright © 2012 · Magnolia Press ISSN 1175-5334 (online edition) A list of Cuban Lepidoptera (Arthropoda: Insecta) RAYNER NÚÑEZ AGUILA1,3 & ALEJANDRO BARRO CAÑAMERO2 1División de Colecciones Zoológicas y Sistemática, Instituto de Ecología y Sistemática, Carretera de Varona km 3. 5, Capdevila, Boyeros, Ciudad de La Habana, Cuba. CP 11900. Habana 19 2Facultad de Biología, Universidad de La Habana, 25 esq. J, Vedado, Plaza de La Revolución, La Habana, Cuba. 3Corresponding author. E-mail: rayner@ecologia. cu Table of contents Abstract . 1 Introduction . 1 Materials and methods. 2 Results and discussion . 2 List of the Lepidoptera of Cuba . 4 Notes . 48 Acknowledgments . 51 References . 51 Appendix . 56 Abstract A total of 1557 species belonging to 56 families of the order Lepidoptera is listed from Cuba, along with the source of each record. Additional literature references treating Cuban Lepidoptera are also provided. The list is based primarily on literature records, although some collections were examined: the Instituto de Ecología y Sistemática collection, Havana, Cuba; the Museo Felipe Poey collection, University of Havana; the Fernando de Zayas private collection, Havana; and the United States National Museum collection, Smithsonian Institution, Washington DC. One family, Schreckensteinidae, and 113 species constitute new records to the Cuban fauna. The following nomenclatural changes are proposed: Paucivena hoffmanni (Koehler 1939) (Psychidae), new comb., and Gonodontodes chionosticta Hampson 1913 (Erebidae), syn. -

Introductions for Biological Control in Hawaii 1997–2001
BPROCIOLOGICAL. HAWAIIAN CONTROL ENTOMOL IN H. SAWAIIOC. (2003), 1997–2001 36:145–153 145 Introductions for Biological Control in Hawaii 1997–2001 Thomas W. Culliney1, Walter T. Nagamine, and Kenneth K. Teramoto Hawaii Department of Agriculture, Division of Plant Industry, Plant Pest Control Branch 1428 South King Street, Honolulu, Hawaii 96814, U.S.A. 1Correspondence: T.W. Culliney, USDA, APHIS, PPQ, Center for Plant Health Science and Technology 1017 Main Campus Dr., Ste. 2500, Raleigh, NC 27606, USA Abstract. Introductions and releases of natural enemies for the biological control of agricultural and forest pests in Hawaii are discussed for the period 1997-2001. Sixteen insect and five fungal species were introduced, released, or redistributed by the Ha- waii Department of Agriculture for the control of six weeds (Clidemia hirta, Coccinia grandis, Miconia calvescens, Myrica faya, Senecio madagascariensis, and Ulex europaeus) and four insect pests (Aleurocanthus woglumi, Bemisia argentifolii, Pentalonia nigronervosa, and Sipha flava). Key words: biological control, Hawaii In its role as the state’s lead agency for classical biological control, the Plant Pest Control Branch of the Hawaii Department of Agriculture (HDOA) has maintained a program for the introduction and release of beneficial organisms for the past 100 years. This paper, the latest in a series, provides information on the status of some pests and their introduced natural enemies, and a list of natural enemies released for biological control in Hawaii from 1997 through 2001 (Table 1). All introductions are thoroughly studied and screened under quar- antine following established protocols, and must be approved by the state Board of Agricul- ture and the United States Department of Agriculture’s (USDA) Animal and Plant Health Inspection Service before they can be released in Hawaii. -

John Pennekamp Coral Reef State Park 2018 Draft Unit Management
John Pennekamp Coral Reef State Park Advisory Group Draft Unit Management Plan STATE OF FLORIDA DEPARTMENT OF ENVIRONMENTAL PROTECTION Division of Recreation and Parks August 2018 TABLE OF CONTENTS INTRODUCTION ...................................................................................1 PURPOSE AND SIGNIFICANCE OF THE PARK ....................................... 3 Park Significance ...............................................................................4 PURPOSE AND SCOPE OF THE PLAN..................................................... 4 MANAGEMENT PROGRAM OVERVIEW ................................................ 10 Management Authority and Responsibility ........................................... 10 Park Management Goals ................................................................... 11 Management Coordination ................................................................ 11 Public Participation ............................................................................ 12 Other Designations ........................................................................... 12 RESOURCE MANAGEMENT COMPONENT INTRODUCTION ................................................................................. 13 RESOURCE DESCRIPTION AND ASSESSMENT .................................... 14 Natural Resources ............................................................................. 14 Topography ................................................................................. 14 Geology ..................................................................................... -

Insecta Mundi 0789: 1–56
InsectaA journal of world insect systematics Mundi 0789 A new genus and 37 new noctuoid species from peninsular Page Count: 56 Florida and the Keys (Lepidoptera: Noctuoidea) J. T. Troubridge 23396 Mullins Ave Port Charlotte, FL, U.S.A. 33954 Date of issue: September 25, 2020 Center for Systematic Entomology, Inc., Gainesville, FL Troubridge JT. 2020. A new genus and 37 new noctuoid species from peninsular Florida and the Keys (Lepidop- tera: Noctuoidea). Insecta Mundi 0789: 1–56. Published on September 25, 2020 by Center for Systematic Entomology, Inc. P.O. Box 141874 Gainesville, FL 32614-1874 USA http://centerforsystematicentomology.org/ Insecta Mundi is a journal primarily devoted to insect systematics, but articles can be published on any non- marine arthropod. Topics considered for publication include systematics, taxonomy, nomenclature, checklists, faunal works, and natural history. Insecta Mundi will not consider works in the applied sciences (i.e. medi- cal entomology, pest control research, etc.), and no longer publishes book reviews or editorials. Insecta Mundi publishes original research or discoveries in an inexpensive and timely manner, distributing them free via open access on the internet on the date of publication. Insecta Mundi is referenced or abstracted by several sources, including the Zoological Record and CAB Abstracts. Insecta Mundi is published irregularly throughout the year, with completed manuscripts assigned an individual number. Manuscripts must be peer reviewed prior to submission, after which they are reviewed by the editorial board to ensure quality. One author of each submitted manuscript must be a current member of the Center for Systematic Entomology. Guidelines and requirements for the preparation of manuscripts are available on the Insecta Mundi website at http://centerforsystematicentomology.org/insectamundi/ Chief Editor: David Plotkin, [email protected] Assistant Editor: Paul E. -

Universityof Hawai" Library Phenology, Reproductive Potential, Seed Dispersal and Predation, and Seedling Establishment Of
4445 UNIVERSITYOF HAWAI" LIBRARY PHENOLOGY, REPRODUCTIVE POTENTIAL, SEED DISPERSAL AND PREDATION, AND SEEDLING ESTABLISHMENT OF THREE INVASIVE PLANT SPECIES IN A HAWAIIAN RAIN FOREST A THESIS SUBMITTED TO THE GRADUATE DIVISION OF THE UNIVERSITY OF HAWAI'I IN PARTIAL FULFILLMENT OF THE REQUIREMENTS FOR THE DEGREE OF DOCTOR OF PHILOSOPHY IN BOTANY MAY 2004 By Arthur C. Medeiros Dissertation Committee: Gerald Carr, Chairperson Sheila Conant David Duffy Lloyd Loope Robert Robichaux DEDICATION PAGE: This work is dedicated to the memory of a brilliant and beloved mentor, Dr. Charles lamoureux, a model teacher, a pioneering scientist in Hawai'i, and a great person. iii ACKNOWLEDGEMENTS This research project was made possible only through support by the Natural Resource Preservation Program (NRPP) of the U.S. Geological Survey (USGS) and support of the Pacific Island Ecosystems Research Center (PIERC) of the USGS. Mahalo loa to Dr. William Steiner and Dr. David Helweg, who head the research center. Mahalo loa to my dissertation committee: Dr. Gerald Carr (chairperson), Dr. Sheila Conant, Dr. David Duffy, Dr. Lloyd Loope, and Dr. Robert Robichaux. My chairperson Dr. Gerald Carr has been a conscientious and warm mentor who has made my graduate experience uncommonly rewarding. Dr. Sheila Conant has always been in my mind a sterling example of what excellence over time is all about. Dr. David Duffy, stepped bravely in to replace committee member Dr. Charles Lamoureux who died unexpectedly in 2001. Dr. Duffy's skills as an editor and his keen intellect have benefited this dissertation substantially. Dr. Lloyd Loope helped develop the initial concepts and scope of the research project which turned out to be my dissertation project. -
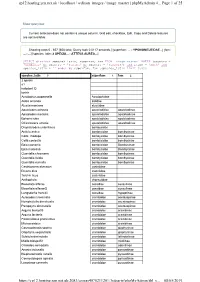
Page 1 of 25 Cp12.Hosting.Zen.Net.Uk / Localhost / Wdixon Images / Image Master | Phpmyadmin 4... 08/05/2019
cp12.hosting.zen.net.uk / localhost / wdixon_images / image_master | phpMyAdmin 4... Page 1 of 25 Show query box Current selection does not contain a unique column. Grid edit, checkbox, Edit, Copy and Delete features are not available. Showing rows 0 - 657 (658 total, Query took 0.0117 seconds.) [superfam: ... - YPONOMEUTIDAE...] [fam: ... - ...] [species_latin: 2 SPECIS... - ATTEVA AUREA...] SELECT distinct species_latin, superfam, fam FROM `image_master` WHERE (country = 'Honduras' or country = 'Panama' or country = 'Pan2018') and itype = 'moth' and species_latin > '' order by superfam, fam ,species_latin limit 0,850 species_latin 3 superfam 1 fam 2 2 specis a1 notodont Q tortrix Acrolophus popeanella Acrolophidae Aidos amanda aididae Alucita montana alucitidae Apatelodes adrastia apatelodidae apatelodinae Apatelodes merlona apatelodidae apatelodinae Ephoria lybia apatelodidae apatelodinae Olceclostera amoria apatelodidae apatelodinae Drepatelodes umbrillinea bombycidae Anticla antica bombycidae bombycinae Colla rhodope bombycidae bombycinae Colla coelestis bombycidae bombycinae Epia casnonia bombycidae Bombycinae Epia muscosa bombycidae Bombycinae Quentalia chromana bombycidae bombycinae Quentalia lividia bombycidae bombycinae Quentalia numalia bombycidae bombycinae Castniomera atymnius castniidae Divana diva castniidae Telchin licus castniidae Anthophyla choreutidae Biocellata alfarae cossidae cossulinae Biocellata alfaraeQ cossidae cossulinae Langsdorfia franckii cossidae hypoptinae Aulacodes traversalis crambidae acentropinae Nymphuliella -

I:\Approved Plans\District 5\Bahia Honda\05-23-2003 Approved Plan
BAHIA HONDA STATE PARK UNIT MANAGEMENT PLAN APPROVED PLAN STATE OF FLORIDA DEPARTMENT OF ENVIRONMENTAL PROTECTION Division of Recreation and Parks MAY 23, 2003 Department of Environmental Protection Jeb Bush Marjorie Stoneman Douglas Building David B. Struhs Governor 3900 Commonwealth Boulevard, MS 140 Secretary Tallahassee, Florida 32399-3000 May 23, 2003 Ms. BryAnne White Government Operations Consultant II Office of Park Planning Division of Recreation and Parks Bahia Honda State Park Lease Number: # 3609 Dear Ms. White: The Division of State Lands has completed the review of Bahia State Park Land Management Plan and find that it fulfills all the requirements of Rule 18-2.021, F.A.C., and ss. 253.034 and 259.032, F.S. Therefore, on May 23, 2003, the Office of Environmental Services, acting as agent for the Board of Trustees of the Internal Improvement Trust Fund approves this plan. The plans five-year update will be due in May 2008. Approval of this land management plan does not waive the authority or jurisdiction of any governmental entity that may have an interest in this project. Implementation of any upland activities proposed by this management plan may require a permit or other authorization from federal and state agencies having regulatory jurisdiction over those particular activities. Sincerely, Delmas T. Barber Delmas T. Barber, OMC Manager Office of Environmental Services Division of State Lands More Protection, Less Process Printed on recycled paper. TABLE OF CONTENTS INTRODUCTION 1 PURPOSE AND SCOPE OF PLAN 1 MANAGEMENT -

RAPPORT Trinité
MISSION ENTOMOLOGIQUE Octobre 2010 Réserve Naturelle de la Trinité, site de Roche Bénitier Etude réalisée par la Etude commandée par Mission entomologique SEAG Octobre 2010 – Réserve Trinité / Station Roche Bénitier Réserve Naturelle de la Trinité Site de Roche Bénitier Mission d’inventaire entomologique Octobre 2010 Rapport Objet : Ce rapport fait état des résultats obtenus suite à l’échantillonnage effectué sur le site de Roche Bénitier dans la Réserve Naturelle de la Trinité par la Société Entomologique Antilles-Guyane du 2 octobre 2010 au 10 octobre 2010. Le rapport présente les différentes méthodes mises en œuvre, quelques photos par famille, des commentaires et quelques travaux statistiques sur les familles et ordres étudiés. Les listes d’espèces déterminées ainsi que les déterminateurs sont donnés en annexe. Maître d’ouvrage : Office National des Forêts Direction Régionale de Guyane Contact : Marguerite Delaval, conservateur de la Réserve de la Trinité Réalisation de l’étude Société entomologique Antilles-Guyane Président : Pierre-Henri DALENS Association Loi 1901 06 94 26 14 76 N° SIRET : 498 671 742 00019 Mail : [email protected] 18 Lotissement Amaryllis Secrétaire : Stéphane BRÛLÉ 97354 Rémire-Montjoly 06 94 26 21 19 Mail : [email protected] Citation conseillée du rapport : BRÛLÉ S., TOUROULT J. & DALENS P.-H., (coord.). 2011. -- Résultats de la mission entomologique du site de Roche Bénitier, Réserve de la Trinité (Guyane), Octobre 2010 . Rapport de la Société entomologique Antilles-Guyane, SEAG, ONF, 93 pp.+ annexes. Non publié. Mission entomologique SEAG Octobre 2010 – Réserve Trinité / Station Roche Bénitier Remerciements Nous tenons à remercier Marguerite DELAVAL (ONF), conservateur de la Réserve Naturelle de la Trinité. -

An Annotated List of the Lepidoptera of Honduras
University of Nebraska - Lincoln DigitalCommons@University of Nebraska - Lincoln Center for Systematic Entomology, Gainesville, Insecta Mundi Florida 2-29-2012 An annotated list of the Lepidoptera of Honduras Jacqueline Y. Miller University of Florida, [email protected] Deborah L. Matthews University of Florida, [email protected] Andrew D. Warren University of Florida, [email protected] M. Alma Solis Systematic Entomology Laboratory, PSI, Agriculture Research Service, USDA, [email protected] Donald J. Harvey Smithsonian Institution, Washington, D.C., [email protected] See next page for additional authors Follow this and additional works at: https://digitalcommons.unl.edu/insectamundi Part of the Entomology Commons Miller, Jacqueline Y.; Matthews, Deborah L.; Warren, Andrew D.; Solis, M. Alma; Harvey, Donald J.; Gentili- Poole, Patricia; Lehman, Robert; Emmel, Thomas C.; and Covell, Charles V., "An annotated list of the Lepidoptera of Honduras" (2012). Insecta Mundi. 725. https://digitalcommons.unl.edu/insectamundi/725 This Article is brought to you for free and open access by the Center for Systematic Entomology, Gainesville, Florida at DigitalCommons@University of Nebraska - Lincoln. It has been accepted for inclusion in Insecta Mundi by an authorized administrator of DigitalCommons@University of Nebraska - Lincoln. Authors Jacqueline Y. Miller, Deborah L. Matthews, Andrew D. Warren, M. Alma Solis, Donald J. Harvey, Patricia Gentili-Poole, Robert Lehman, Thomas C. Emmel, and Charles V. Covell This article is available at DigitalCommons@University of Nebraska - Lincoln: https://digitalcommons.unl.edu/ insectamundi/725 INSECTA A Journal of World Insect Systematics MUNDI 0205 An annotated list of the Lepidoptera of Honduras Jacqueline Y. Miller, Deborah L.