Flora of Whiteknights Park
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

"Ecology of Water Relations in Plants". In: Encyclopedia of Life
Ecology of Water Relations Advanced article in Plants Article Contents . Introduction Yoseph Negusse Araya, The Open University, Milton Keynes, UK . Water Uptake and Movement through Plants . Water Stress and Plants Water is an important resource for plant growth. Availability of water in the soil determines . Plant Sensing and Adaptation to Water Stress the niche, distribution and competitive interaction of plants in the environment. Distribution of Plants in Response to Water Regime Introduction doi: 10.1002/9780470015902.a0003201 Importance of water for plants Moisture Water typically constitutes 80–95% of the mass of growing 8 plant tissues and plays a crucial role for plant growth (Taiz 7 and Zeiger, 1998). Plants require water for a number of 6 physiological processes (e.g. synthesis of carbohydrates) 5 and for associated physical functions (e.g. keeping plants turgid). 4 Water accomplishes its many functions because of its 3 Moisture index 2 unique characteristics: the polarity of the molecule H2O (which makes it an excellent solvent), viscosity (which 1 makes it capable of moving through plant tissues by 0 capillary action) and thermal properties (which makes it Forest Woodland Grassland Desert capable of cooling plant tissues). Total net productivity 1 Plants require water, soil nutrients, carbon dioxide, ox- − 1000 year ygen and solar radiation for growth. Of these, water is most 2 − often the most limiting: influencing productivity (Taiz and m 1 800 Zeiger, 1998) as well as the diversity of species (Rodriguez- − Iturbe and Porporato, 2004) in both natural and agricul- 600 tural ecosystems. This is illustrated graphically in Figure 1. 400 How does water affect ecology of plants? 200 In order to understand the ecology of plant–water rela- 0 Total net productivity g Total tions it is important to understand from where and how Forest Woodland Grassland Desert plants acquire water in their environment (the latter is dis- cussed in the section on water uptake and movement Plant species diversity through plants). -

Generative Reproduction of Antarctic Grasses, the Native Species Deschampsia Antarctica Desv
vol. 36, no. 3, pp. 261–279, 2015 doi: 10.1515/popore−2015−0016 Generative reproduction of Antarctic grasses, the native species Deschampsia antarctica Desv. and the alien species Poa annua L. Irena GIEŁWANOWSKA1,2* and Wioleta KELLMANN−SOPYŁA1 1Katedra Fizjologii, Genetyki i Biotechnologii Roślin, Wydział Biologii i Biotechnologii, Uniwersytet Warmińsko−Mazurski w Olsztynie, ul. Oczapowskiego 1A, 10−719 Olsztyn, Poland 2 Instytut Biochemii i Biofizyki PAN, Zakład Biologii Antarktyki i Polska Stacja Antarktyczna “H. Arctowski”, ul. Ustrzycka 10/12, 02−141 Warszawa, Poland * corresponding author <[email protected]> Abstract: The embryology of two species, Deschampsia antarctica, a native species, and Poa annua, an alien species in the Antarctic we studied. Flowering buds of plants growing in their natural habitats on King George Island and generative tissues of both plant species grown in a greenhouse were analyzed. Adaptations to autogamy and anemogamy were ob− served in the flower anatomy of both species. The microsporangia of the evaluated grasses produce a small number of three−celled pollen grains. Numerous pollen grains do not leave the microsporangium and germinate in the thecae. Deschampsia antarctica and P. annua plants harvested in Antarctica developed a particularly small number of microspores in pol− len chambers. In D. antarctica, male gametophytes were produced at a faster rate: genera− tive cells in pollen did not become detached from the wall of the pollen grain, they were not embedded in the cytoplasm of vegetative cells, and they divided into two sperm cells situ− ated close to the wall. The monosporous Polygonum type of embryo sac development was observed in the studied species. -

Breeding System Diversification and Evolution in American Poa Supersect. Homalopoa (Poaceae: Poeae: Poinae)
Annals of Botany Page 1 of 23 doi:10.1093/aob/mcw108, available online at www.aob.oxfordjournals.org Breeding system diversification and evolution in American Poa supersect. Homalopoa (Poaceae: Poeae: Poinae) Liliana M. Giussani1,*, Lynn J. Gillespie2, M. Amalia Scataglini1,Marıa A. Negritto3, Ana M. Anton4 and Robert J. Soreng5 1Instituto de Botanica Darwinion, San Isidro, Buenos Aires, Argentina, 2Research and Collections Division, Canadian Museum of Nature, Ottawa, Ontario, Canada, 3Universidad de Magdalena, Santa Marta, Colombia, 4Instituto Multidisciplinario de Biologıa Vegetal (IMBIV), CONICET-UNC, Cordoba, Argentina and 5Department of Botany, Smithsonian Institution, Washington, DC, USA *For correspondence. E-mail [email protected] Received: 11 December 2015 Returned for revision: 18 February 2016 Accepted: 18 March 2016 Downloaded from Background and Aims Poa subgenus Poa supersect. Homalopoa has diversified extensively in the Americas. Over half of the species in the supersection are diclinous; most of these are from the New World, while a few are from South-East Asia. Diclinism in Homalopoa can be divided into three main types: gynomonoecism, gynodioe- cism and dioecism. Here the sampling of species of New World Homalopoa is expanded to date its origin and diver- sification in North and South America and examine the evolution and origin of the breeding system diversity. Methods A total of 124 specimens were included in the matrix, of which 89 are species of Poa supersect. http://aob.oxfordjournals.org/ Homalopoa sections Acutifoliae, Anthochloa, Brizoides, Dasypoa, Dioicopoa, Dissanthelium, Homalopoa sensu lato (s.l.), Madropoa and Tovarochloa, and the informal Punapoa group. Bayesian and parsimony analyses were conducted on the data sets based on four markers: the nuclear ribosomal internal tanscribed spacer (ITS) and exter- nal transcribed spacer (ETS), and plastid trnT-L and trnL-F. -

Anthracene Derivatives in Some Species of Rumex L
Vol. 76, No. 2: 103-108, 2007 ACTA SOCIETATIS BOTANICORUM POLONIAE 103 ANTHRACENE DERIVATIVES IN SOME SPECIES OF RUMEX L. GENUS MAGDALENA WEGIERA1, HELENA D. SMOLARZ1, DOROTA WIANOWSKA2, ANDRZEJ L. DAWIDOWICZ2 1 Departament of Pharmaceutical Botany Skubiszewski Medical University of Lublin Chodki 1, 20-039 Lublin, Poland e-mail: [email protected] 2 Faculty of Chemistry, Marii Curie-Sk³odowska University of Lublin (Received: June 1, 2006. Accepted: September 5, 2006) ABSTRACT Eight anthracene derivatives (chrysophanol, physcion, emodin, aloe-emodin, rhein, barbaloin, sennoside A and sennoside B) were signified in six species of Rumex L genus: R. acetosa L., R. acetosella L., R. confertus Willd., R. crispus L., R. hydrolapathum Huds. and R. obtusifolius L. For the investigations methanolic extracts were pre- pared from the roots, leaves and fruits of these species. Reverse Phase High Performance Liquid Chromatography was applied for separation, identification and quantitative determination of anthracene derivatives. The identity of these compounds was further confirmed with UV-VIS. Received data were compared. The roots are the best organs for the accumulation of anthraquinones. The total amount of the detected compo- unds was the largest in the roots of R. confertus (163.42 mg/g), smaller in roots R. crispus (25.22 mg/g) and the smallest in roots of R. hydrolapathum (1.02 mg/g). KEY WORDS: Rumex sp., roots, fruits, leaves, anthracene derivatives, RP-HPLC. INTRODUCTION scion-anthrone, rhein, nepodin, nepodin-O-b-D-glycoside and 1,8-dihydroxyanthraquinone from R. acetosa (Dedio The species belonging to the Rumex L. genus are wide- 1973; Demirezer and Kuruuzum 1997; Fairbairn and El- spread in the world. -

Research Regarding the Melliferous Charactheristics of Labiates from Xerophile Meadows from Danube Valley
Lucr ări ştiin Ńifice Zootehnie şi Biotehnologii , vol. 41(1) (2008 ), Timi şoara RESEARCH REGARDING THE MELLIFEROUS CHARACTHERISTICS OF LABIATES FROM XEROPHILE MEADOWS FROM DANUBE VALLEY CERCET ĂRI ASUPRA VALORII MELIFERE A LAMIACEELOR DIN PAJI ŞTILE XEROFITE DIN LUNCA DUN ĂRII ION NICOLETA Apiculture Research and Development Institute of Bucharest, Romania The xerophile meadows in the Danube Valley are dry meadows, located at a great distance from the Danube and with underground waters at greater depth. Their floral composition is characterized by a small number of species pertaining to both mezoxerophiles and to xerophiles, yet the highest percentage is covered by xerophile species, which are characterized by their small foliage surface, the very narrow and tough limb, and acute porosity etc.In the floral composition of these species, the graminaceae species are best represented, followed by leguminous and lamiaceae, known in beekeeping as good honey plants. Thus, the researches carried out have shown that Lamiaceae species have a good participation, with variation limits raging from 15% to 50-60%. Leguminous species are represented less on xerophile meadows than in hidrophile meadows. Among these we mention: Lotus corniculatus L., Trifolium repens L. şi Medicago lupulina L., all these species being known in beekeeping as good honey plants. Among gramineae species the most representatives are: Lolium perene L. and Poa pratensis L., yet with no melliferous value. Likewise, the group of „various” plants varied a lot as participation in the structure of the vegetal cover of xerophile meadows, depending on the place of research, all these species having no melliferous value. The current paper describes the results o biometric and melliferous researches carried out over the period 2003- 2005 on 5 plant species pertaining to the Lamiaceae family, namely: Salvia nemerosa L. -

Towards Resolving Lamiales Relationships
Schäferhoff et al. BMC Evolutionary Biology 2010, 10:352 http://www.biomedcentral.com/1471-2148/10/352 RESEARCH ARTICLE Open Access Towards resolving Lamiales relationships: insights from rapidly evolving chloroplast sequences Bastian Schäferhoff1*, Andreas Fleischmann2, Eberhard Fischer3, Dirk C Albach4, Thomas Borsch5, Günther Heubl2, Kai F Müller1 Abstract Background: In the large angiosperm order Lamiales, a diverse array of highly specialized life strategies such as carnivory, parasitism, epiphytism, and desiccation tolerance occur, and some lineages possess drastically accelerated DNA substitutional rates or miniaturized genomes. However, understanding the evolution of these phenomena in the order, and clarifying borders of and relationships among lamialean families, has been hindered by largely unresolved trees in the past. Results: Our analysis of the rapidly evolving trnK/matK, trnL-F and rps16 chloroplast regions enabled us to infer more precise phylogenetic hypotheses for the Lamiales. Relationships among the nine first-branching families in the Lamiales tree are now resolved with very strong support. Subsequent to Plocospermataceae, a clade consisting of Carlemanniaceae plus Oleaceae branches, followed by Tetrachondraceae and a newly inferred clade composed of Gesneriaceae plus Calceolariaceae, which is also supported by morphological characters. Plantaginaceae (incl. Gratioleae) and Scrophulariaceae are well separated in the backbone grade; Lamiaceae and Verbenaceae appear in distant clades, while the recently described Linderniaceae are confirmed to be monophyletic and in an isolated position. Conclusions: Confidence about deep nodes of the Lamiales tree is an important step towards understanding the evolutionary diversification of a major clade of flowering plants. The degree of resolution obtained here now provides a first opportunity to discuss the evolution of morphological and biochemical traits in Lamiales. -

Concentrations of Anthraquinone Glycosides of Rumex Crispus During Different Vegetation Stages L
Concentrations of Anthraquinone Glycosides of Rumex crispus during Different Vegetation Stages L. Ömtir Demirezer Hacettepe University, Faculty of Pharmacy, Department of Pharmacognosy, 06100 Ankara, Turkey Z. Naturforsch. 49c, 404-406 (1994); received January 31, 1994 Rumex crispus, Polygonaceae. Anthraquinone, Glycoside The anthraquinone glycoside contents of various parts of Rumex crispus L. (Polygonaceae) in different vegetation stages were investigated by thin layer chromatographic and spectro- photometric methods. The data showed that the percentage of anthraquinone glycoside in all parts of plant increased at each stage. Anthraquinone glycoside content was increased in leaf, stem, fruit and root from 0.05 to 0.40%. from 0.03 to 0.46%. from 0.08 to 0.34%, and from 0.35 to 0.91% respectively. From the roots of R. crispus, emodin- 8 -glucoside, RGA (isolated in our laboratory, its structure was not elucidated), traceable amount of glucofran- gulin B and an unknown glycoside ( R f = 0.28 in ethyl acetate:methanol:water/100:20:10) was detected in which the concentration was increased from May to August. The other parts of plant contained only emodin- 8 -glucoside. Introduction In the present investigation various parts of Rumex L. (Polygonaceae) is one of several Rumex crispus, leaf, stem, fruit and root were ana genera which is characterized by the presence lyzed separately for their anthraquinone glycoside of anthraquinone derivatives. There are about contents, the glycosides in different vegetation 200 species of Rumex in worldwide (Hegi, 1957). stages were detected individually. By this method, Rumex is represented with 23 species and 5 hy translocation of anthraquinone glycosides were brids in Turkey (Davis, 1965) and their roots have also investigated. -

Fort Ord Natural Reserve Plant List
UCSC Fort Ord Natural Reserve Plants Below is the most recently updated plant list for UCSC Fort Ord Natural Reserve. * non-native taxon ? presence in question Listed Species Information: CNPS Listed - as designated by the California Rare Plant Ranks (formerly known as CNPS Lists). More information at http://www.cnps.org/cnps/rareplants/ranking.php Cal IPC Listed - an inventory that categorizes exotic and invasive plants as High, Moderate, or Limited, reflecting the level of each species' negative ecological impact in California. More information at http://www.cal-ipc.org More information about Federal and State threatened and endangered species listings can be found at https://www.fws.gov/endangered/ (US) and http://www.dfg.ca.gov/wildlife/nongame/ t_e_spp/ (CA). FAMILY NAME SCIENTIFIC NAME COMMON NAME LISTED Ferns AZOLLACEAE - Mosquito Fern American water fern, mosquito fern, Family Azolla filiculoides ? Mosquito fern, Pacific mosquitofern DENNSTAEDTIACEAE - Bracken Hairy brackenfern, Western bracken Family Pteridium aquilinum var. pubescens fern DRYOPTERIDACEAE - Shield or California wood fern, Coastal wood wood fern family Dryopteris arguta fern, Shield fern Common horsetail rush, Common horsetail, field horsetail, Field EQUISETACEAE - Horsetail Family Equisetum arvense horsetail Equisetum telmateia ssp. braunii Giant horse tail, Giant horsetail Pentagramma triangularis ssp. PTERIDACEAE - Brake Family triangularis Gold back fern Gymnosperms CUPRESSACEAE - Cypress Family Hesperocyparis macrocarpa Monterey cypress CNPS - 1B.2, Cal IPC -

A List of Grasses and Grasslike Plants of the Oak Openings, Lucas County
A LIST OF THE GRASSES AND GRASSLIKE PLANTS OF THE OAK OPENINGS, LUCAS COUNTY, OHIO1 NATHAN WILLIAM EASTERLY Department of Biology, Bowling Green State University, Bowling Green, Ohio 4-3403 ABSTRACT This report is the second of a series of articles to be prepared as a second "Flora of the Oak Openings." The study represents a comprehensive survey of members of the Cyperaceae, Gramineae, Juncaceae, Sparganiaceae, and Xyridaceae in the Oak Openings region. Of the 202 species listed in this study, 34 species reported by Moseley in 1928 were not found during the present investigation. Fifty-seven species found by the present investi- gator were not observed or reported by Moseley. Many of these species or varieties are rare and do not represent a stable part of the flora. Changes in species present or in fre- quency of occurrence of species collected by both Moseley and Easterly may be explained mainly by the alteration of habitats as the Oak Openings region becomes increasingly urbanized or suburbanized. Some species have increased in frequency on the floodplain of Swan Creek, in wet ditches and on the banks of the Norfolk and Western Railroad right-of-way, along newly constructed roadsides, or on dry sandy sites. INTRODUCTION The grass family ranks third among the large plant families of the world. The family ranks number one as far as total numbers of plants that cover fields, mead- ows, or roadsides are concerned. No other family is used as extensively to pro- vide food or shelter or to create a beautiful landscape. The sedge family does not fare as well in terms of commercial importance, but the sedges do make avail- able forage and food for wild fowl and they do contribute plant cover in wet areas where other plants would not be as well adapted. -

Invasive Alien Species in Southern Africa
GISP Global Invasive Species Programme Ministry of Tourism, Environment United States Government and Natural Resources Republic of Zambia Invasive Alien Species in Southern Africa National Reports & Directory of Resources Edited by Ian A.W. Macdonald, Jamie K. Reaser, Chris Bright, Laurie E. Neville, Geoffrey W. Howard, Sean J. Murphy, and Guy Preston 1 This report is a product of a workshop entitled Prevention and Management of Invasive Alien Species: Forging Cooperation throughout Southern Africa, held by the Global Invasive Species Programme (GISP) in Lusaka, Zambia on 10-12 June 2002. It was sponsored by the U.S. Department of State, Bureau of Oceans and International Environmental Affairs (OESI) grant S-LMAQM-00-H-0167. In-kind assistance was provided by the U.S. Environmental Protection Agency. Administrative and logistical assistance was provided by the Zambian Ministry of Tourism, Environment and Natural Resources, the U.S. Embassy in Lusaka, Zambia, the Scientific Committee on Problems of the Environment (SCOPE), and the National Fish and Wildlife Foundation (NFWF), as well as all Steering Committee members. The Smithsonian Institution National Museum of Natural History and National Botanical Institute, South Africa kindly provided support during report production. The editors thank Dr Phoebe Barnard of the GISP Secretariat for her very extensive work to finalize the report. The workshop was co-chaired by the Governments of the Republic of Zambia and the United States of America, and by the Global Invasive Species Programme. Members of the Steering Committee included: Mr Lubinda Aongola (Ministry of Tourism, Environment and Natural Resources, Zambia), Mr Troy Fitrell (U.S. -
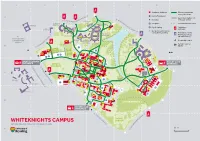
Whiteknights-Campus-Map.Pdf
1 2 3 4 5 6 7 Academic buildings Primary pedestrain and cycle routes Halls of residence A D A Secondary pedestrain RO S W Bus stop and cycle routes UPP ND H ER RED L A BROADOAK IT PLACE E K DUNSDEN N 1 Car parks Unsurfaced/unlit paths I CRESCENT G WANTAGE H CHILDS T S £ Pay & display Pedestrian ST R entrance GEORGES 84 OA D Car parks restricted to users of this building Pedestrian exit to STENTON Wokingham Road MACKINDER TOWNHOUSES D off-site housing A 202 TOWN CENTRE & O R WINDSOR, Accessible toilets LONDON ROAD T 295 PARK S EAT GREENOW & CAMPUS R U MCCOMBIE H STENTON Gender-neutral M TENNIS WESSEX B EL PARK toilets 2 253 3 34 39 71 16 1a £ 50 1 £ 30 BRIDGES 53 76 138 196 21 SHINFIELD ROAD 195 EARLEY GATE ENTRANCE SPORTS 26 ENTRANCE 9 135 4 GROUNDS 1 NORTHCOURT AVENUE S 5 H 29 54 I N 201 C F 27 I E L 55 D 37 8 R 259 7 20 O FRIENDS A 2 BRIDGE 56 BENYON D 6 33 15 9 21 7 70 40 ST PATRICKS 10 49 48 25 3 118 62 5 24 22 74 10 6 59 SHERFIELD 4 58 8 46 23 38 47 261 D 32 13 31 THE WILDERNESS 11 210 PEPPER LANE 186 ENTRANCE P E P D P A E O R R L HARRIS S A S N GARDEN E WHITEKNIGHTS CAMPUS E N R E SAT NAV RG6 6UR | RG6 7BE FOR EARLEY GATE D IL W E N 200M WHITEKNIGHTS MAP KEY (A–Z) Name and grid ref Building Name and grid ref Building Name and grid ref Building Name and grid ref Building 29 3sixty bar C4 Students’ Union 49 Department of Languages Miller 135 Job Shop (students) C4 Carrington 62 Speech and Language Therapy Psychology and Cultures C3 Clinic D6 135 Accommodation Office C4 Carrington 38 Knowledge Transfer Centre D4 Polly -

Georg-August-Universität Göttingen
GÖTTINGER ZENTRUM FÜR BIODIVERSITÄTSFORSCHUNG UND ÖKOLOGIE GÖTTINGEN CENTRE FOR BIODIVERSITY AND ECOLOGY Herb layer characteristics, fly communities and trophic interactions along a gradient of tree and herb diversity in a temperate deciduous forest Dissertation zur Erlangung des Doktorgrades der Mathematisch-Naturwissenschaftlichen Fakultäten der Georg-August-Universität Göttingen vorgelegt von Mag. rer. nat. Elke Andrea Vockenhuber aus Wien Göttingen, Juli, 2011 Referent: Prof. Dr. Teja Tscharntke Korreferent: Prof. Dr. Stefan Vidal Tag der mündlichen Prüfung: 16.08.2011 2 CONTENTS Chapter 1: General Introduction............................................................................................ 5 Effects of plant diversity on ecosystem functioning and higher trophic levels ....................................................... 6 Study objectives and chapter outline ...................................................................................................................... 8 Study site and study design ................................................................................................................................... 11 Major hypotheses.................................................................................................................................................. 12 References............................................................................................................................................................. 13 Chapter 2: Tree diversity and environmental context