Prognosis Value of RBBP8 Expression in Plasma Cell Myeloma
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Gene Expression Signature–Based Prognostic Risk Score in Patients with Primary Central Nervous System Lymphoma
Published OnlineFirst August 20, 2012; DOI: 10.1158/1078-0432.CCR-12-0596 Clinical Cancer Imaging, Diagnosis, Prognosis Research Gene Expression Signature–Based Prognostic Risk Score in Patients with Primary Central Nervous System Lymphoma Atsushi Kawaguchi1, Yasuo Iwadate2, Yoshihiro Komohara3, Masakazu Sano4, Koji Kajiwara5, Naoki Yajima4, Naoto Tsuchiya4, Jumpei Homma4, Hiroshi Aoki4, Tsutomu Kobayashi4, Yuko Sakai7, Hiroaki Hondoh6, Yukihiko Fujii4, Tatsuyuki Kakuma1, and Ryuya Yamanaka7 Abstract Purpose: Better understanding of the underlying biology of primary central nervous system lymphomas (PCNSL) is critical for the development of early detection strategies, molecular markers, and new therapeutics. This study aimed to define genes associated with survival of patients with PCNSL. Experimental Design: Expression profiling was conducted on 32 PCNSLs. A gene classifier was developed using the random survival forests model. On the basis of this, prognosis prediction score (PPS) using immunohistochemical analysis is also developed and validated in another data set with 43 PCNSLs. Results: We identified 23 genes in which expressions were strongly and consistently related to patient survival. A PPS was developed for overall survival (OS) using a univariate Cox model. Survival analyses using the selected 23-gene classifiers revealed a prognostic value for high-dose methotrexate (HD-MTX) and HD- MTX–containing polychemotherapy regimen–treated patients. Patients predicted to have good outcomes by the PPS showed significantly longer survival than those with poor predicted outcomes (P < 0.0001). PPS using immunohistochemical analysis is also significant in test (P ¼ 0.0004) and validation data set (P ¼ 0.0281). The gene-based predictor was an independent prognostic factor in a multivariate model that included clinical risk stratification (P < 0.0001). -

Prognosis Value of RBBP8 Expression in Plasma Cell Myeloma
Cancer Gene Therapy https://doi.org/10.1038/s41417-018-0069-3 ARTICLE Prognosis value of RBBP8 expression in plasma cell myeloma 1 2 3 4 5 1 1 1 Weilong Zhang ● Ying Song ● Xue He ● Xiaoni Liu ● Ye Zhang ● Zuozhen Yang ● Ping Yang ● Jing Wang ● 1 4 3 4 1 Kai Hu ● Weiyou Liu ● Xiuru Zhang ● Xiaoliang Yuan ● Hongmei Jing Received: 30 July 2018 / Revised: 30 October 2018 / Accepted: 2 November 2018 © The Author(s) 2019. This article is published with open access Abstract Plasma cell myeloma (PCM) secretes monoclonal immunoglobulin (Ig) by clonal plasma cells of abnormal proliferation in the bone marrow. As PCM is incurable, it is necessary to find new biomarkers to predict the prognosis and recurrence of PCM. The relationship between cancer and RBBP8 has not been fully studied. The role of RBBP8 in tumorigenesis remains inconsistent. We described the expression of RBBP8 in the gene expression profile of 1930 PCM samples (1878 PCM patients) from seven independent data sets. We analyzed the relationship between RBBP8 and survival prognosis, recurrence, and treatment response in patients with PCM, and the biological significance of RBBP8 in PCM. The gene expression level of RBBP8 was significantly related to the International staging system (ISS) grade of PCM (P = 0.0012). RBBP8 expression in different molecular subtypes was different (P < 2.2e-16). High RBBP8 expression is associated with 1234567890();,: 1234567890();,: poor survival in PCM (P < 0.0001). High expression of RBBP8 indicates that PCM patients are more likely to relapse (P = 0.0078). The biological significance of RBBP8 in PCM is related to the cell cycle (P < 0.05). -
HCC and Cancer Mutated Genes Summarized in the Literature Gene Symbol Gene Name References*
HCC and cancer mutated genes summarized in the literature Gene symbol Gene name References* A2M Alpha-2-macroglobulin (4) ABL1 c-abl oncogene 1, receptor tyrosine kinase (4,5,22) ACBD7 Acyl-Coenzyme A binding domain containing 7 (23) ACTL6A Actin-like 6A (4,5) ACTL6B Actin-like 6B (4) ACVR1B Activin A receptor, type IB (21,22) ACVR2A Activin A receptor, type IIA (4,21) ADAM10 ADAM metallopeptidase domain 10 (5) ADAMTS9 ADAM metallopeptidase with thrombospondin type 1 motif, 9 (4) ADCY2 Adenylate cyclase 2 (brain) (26) AJUBA Ajuba LIM protein (21) AKAP9 A kinase (PRKA) anchor protein (yotiao) 9 (4) Akt AKT serine/threonine kinase (28) AKT1 v-akt murine thymoma viral oncogene homolog 1 (5,21,22) AKT2 v-akt murine thymoma viral oncogene homolog 2 (4) ALB Albumin (4) ALK Anaplastic lymphoma receptor tyrosine kinase (22) AMPH Amphiphysin (24) ANK3 Ankyrin 3, node of Ranvier (ankyrin G) (4) ANKRD12 Ankyrin repeat domain 12 (4) ANO1 Anoctamin 1, calcium activated chloride channel (4) APC Adenomatous polyposis coli (4,5,21,22,25,28) APOB Apolipoprotein B [including Ag(x) antigen] (4) AR Androgen receptor (5,21-23) ARAP1 ArfGAP with RhoGAP domain, ankyrin repeat and PH domain 1 (4) ARHGAP35 Rho GTPase activating protein 35 (21) ARID1A AT rich interactive domain 1A (SWI-like) (4,5,21,22,24,25,27,28) ARID1B AT rich interactive domain 1B (SWI1-like) (4,5,22) ARID2 AT rich interactive domain 2 (ARID, RFX-like) (4,5,22,24,25,27,28) ARID4A AT rich interactive domain 4A (RBP1-like) (28) ARID5B AT rich interactive domain 5B (MRF1-like) (21) ASPM Asp (abnormal -

Table S1 Primers Used for Genomic DNA Sequencing of K-Ras and TP53
Table S1 Primers used for genomic DNA sequencing of K-Ras and TP53 Gene, exon Forward primer Reverse primer k-Ras, exon 1 GTTCTAATATAGTCACATTT ACTCATGAAAATGGTCAGAGAAACCTTTAT k-Ras, exon 2 GAAGTAAAAGGTGCACTGTA AACTATAATTACTCCTTAAT TP53, exon 4 AGGACCTGGTCCTCTGAC CTGGGAAGGGACAGAAGA TP53, exon 5 TGACTTTCAACTCTGTCTCCT TCAGTGAGGAATCAGAGGCC TP53, exon 6 CTGGAGAGACGACAGAGGCC CCAGAGACCCCAGTTGCAAAC TP53, exon 7 AAGGCGCACTGGCCTCATCTT CGCCGGAAATGTGATGAGAG TP53, exon 8 TGGTTGGGAGTAGATGGAGCC CACCGCTTCTTGTCCTGCTT TP53, exon 9 GTGGAGGAGACCAAGGGTGCA AGGTAAAACAGTCAAGAAGAA Table S2. Genes in cytogenetic band with recurrent copy number alterations Gains Cytoband Gene Protein name Cytoband Gene Protein name 1 q42.2 KIAA1383 KIAA1383 18 q11.1-11.2 KIAA1772 KIAA1772 establishment of cohesion 1 homolog 1 (S. 1 q42.2 C1orf57 chromosome 1 open reading frame 57 18 q11.1-11.2 ESCO1 cerevisiae) 1 q42.2 PCNXL2 pecanex-like 2 (Drosophila) 18 q11.1-11.2 SNRPD1 small nuclear ribonucleoprotein D1 polypeptide 1 q42.2 KIAA1804 mixed lineage kinase 4 18 q11.1-11.2 ABHD3 abhydrolase domain containing 3 11 q13.1 SLC25A45 solute carrier family 25, member 45 18 q11.1-11.2 MIB1 mindbomb homolog 1 (Drosophila) 11 q13.1 FRMD8 FERM domain containing 8 18 q11.1-11.2 GATA6 GATA binding protein 6 11 q13.1 NCRNA00084 non-protein coding RNA 84 18 q11.1-11.2 CTAGE1 cutaneous T-cell lymphoma-associated antigen 1 metastasis associated lung adenocarcinoma 11 q13.1 MALAT1 18 q11.1-11.2 RBBP8 retinoblastoma binding protein 8 transcript 1 (non-protein coding) 20 q13.13 CEBPB CCAAT/enhancer -

Supplemental Material.Pdf
Supplemental Material ZNF750 Interacts with KLF4 and RCOR1, KDM1A, and CTBP1/2 Chromatin Regulators to Repress Epidermal Progenitor Genes and Induce Differentiation Genes Lisa D. Boxer, Brook Barajas, Shiying Tao, Jiajing Zhang, and Paul Khavari Supplemental Inventory Figure S1. This figure supports Figure 1 and shows the Gene Ontology terms for ZNF750-bound but unaffected genes, the genomic enrichment of ZNF750 ChIP-seq peaks, and the percentage of peaks that contain the ZNF750 motif. Figure S2. This figure supports Figure 2 and shows gene expression changes with depletion of ZNF750-interacting proteins, confirms the knock-down of ZNF750-interacting proteins, and shows the quantification of Ki67 in ZNF750-interacting protein depleted organotypic tissue. Figure S3. This figure supports Figure 3 and shows the quantification of ZNF750 and interacting protein co-IPs, and IPs and Far western blots demonstrating competition between KLF4 and KDM1A for binding to ZNF750. Figure S4. This figure supports Figure 4 and shows the expression of ZNF750 mutant proteins and the effects of full-length or mutant ZNF750 on differentiation in organotypic tissue and on clonogenic growth. Figure S5. This figure supports Figure 5 and shows the effects of mutagenesis of ZNF750 and KLF4 motifs on reporter activity, and the changes in histone marks with depletion of ZNF750 and interacting proteins. Figure S6. This figure supports Figure 6 and shows the changes in mRNA and protein expression of ZNF750-interacting proteins during keratinocyte differentiation, and the expression of ZNF750-interacting proteins with ZNF750 depletion. Table S1. Supports Figure 1 and shows the genomic coordinates of ZNF750 ChIP-seq peaks. -

Aberrant Recombination and Repair During Immunoglobulin Class Switching in BRCA1-Deficient Human B Cells
Aberrant recombination and repair during immunoglobulin class switching in BRCA1-deficient human B cells Andrea Björkmana, Per Qvistb,c, Likun Dua, Margarita Bartisha, Apostolos Zaravinosa, Konstantinos Georgioua, Anders D. Børglumb,c, Richard A. Gattid,e, Therese Törngrenf, and Qiang Pan-Hammarströma,1 aDepartment of Laboratory Medicine, Karolinska Institutet, 141 86 Stockholm, Sweden; bDepartment of Biomedicine and cCentre for Integrative Sequencing, Aarhus University, 8000 Aarhus, Denmark; Departments of dPathology and Laboratory Medicine and eHuman Genetics, University of California, Los Angeles, CA 90095; and fDivision of Oncology and Pathology, Department of Clinical Sciences, Lund University, Lund 22100, Sweden Edited by Tak W. Mak, The Campbell Family Institute for Breast Cancer Research at Princess Margaret Cancer Centre, Ontario Cancer Institute, University Health Network, Toronto, Canada, and approved January 7, 2015 (received for review October 3, 2014) Breast cancer type 1 susceptibility protein (BRCA1) has a multitude (1). When the c-NHEJ pathway is defective, alternative end- of functions that contribute to genome integrity and tumor sup- joining (A-EJ) pathway(s), often associated with resections/dele- pression. Its participation in the repair of DNA double-strand tions and longer MHs, may be operating (1–3). breaks (DSBs) during homologous recombination (HR) is well rec- DSBs are not always pathological, but can also be interme- ognized, whereas its involvement in the second major DSB repair diates of physiological processes, such as those that occur during pathway, nonhomologous end-joining (NHEJ), remains controver- B-cell development. Then, extensive gene rearrangements/mod- sial. Here we have studied the role of BRCA1 in the repair of ifications at the Ig gene loci may occur, resulting in production of DSBs in switch (S) regions during immunoglobulin class switch re- functional antibodies that can recognize and act against an im- combination, a physiological, deletion/recombination process that mense number of different pathogens. -

Homologous Repair Deficiency Score for Identifying Breast Cancers With
www.nature.com/scientificreports OPEN Homologous repair defciency score for identifying breast cancers with defective DNA damage response Ahrum Min1,2,9, Kwangsoo Kim1,9, Kyeonghun Jeong1, Seongmin Choi1, Seongyeong Kim2, Koung Jin Suh2,3, Kyung‑Hun Lee2,4*, Sun Kim5,6,7 & Seock‑Ah Im1,2,4,8* Breast cancer (BC) in patients with germline mutations of BRCA1/BRCA2 are associated with beneft from drugs targeting DNA damage response (DDR), but they account for only 5–7% of overall breast cancer. To defne the characteristics of these tumors and also to identify tumors without BRCA mutation but with homologous recombination defciency (HRD) is clinically relevant. To defne characteristic features of HRD tumors and analyze the correlations between BRCA1/BRCA2 and BC subtypes, we analyzed 981 breast tumors from the TCGA database using the signature analyzer. The BRCA signature was strongly associated with the HRD score top 10% (score ≥ 57) population. This population showed a high level of mutations in DDR genes, including BRCA1/BRCA2. HRD tumors were associated with high expression levels of BARD1 and BRIP1. Besides, BRCA1/2 mutations were dominantly observed in basal and luminal subtypes, respectively. A comparison of HRD features in BC revealed that BRCA1 exerts a stronger infuence inducing HRD features than BRCA2 does. It reveals genetic diferences between BRCA1 and BRCA2 and provides a basis for the identifcation of HRD and other BRCA‑associated tumors. Depending on hormone receptor and human epidermal growth factor type II receptor (HER2) oncoprotein expression, breast cancer (BC) is traditionally classifed into luminal A or B (i.e., estrogen and/or progesterone receptor-positive), HER2-enriched, or triple-negative BC (TNBC). -

University of Birmingham Germline RBBP8 Variants Associated With
University of Birmingham Germline RBBP8 variants associated with early- onset breast cancer compromise replication fork stability The COMPLEXO Network DOI: 10.1172/JCI127521 License: None: All rights reserved Document Version Peer reviewed version Citation for published version (Harvard): The COMPLEXO Network 2020, 'Germline RBBP8 variants associated with early-onset breast cancer compromise replication fork stability', Journal of Clinical Investigation, vol. 130, no. 8, pp. 4069-4080. https://doi.org/10.1172/JCI127521 Link to publication on Research at Birmingham portal Publisher Rights Statement: https://doi.org/10.1172/JCI127521. Copyright © 2020, American Society for Clinical Investigation General rights Unless a licence is specified above, all rights (including copyright and moral rights) in this document are retained by the authors and/or the copyright holders. The express permission of the copyright holder must be obtained for any use of this material other than for purposes permitted by law. •Users may freely distribute the URL that is used to identify this publication. •Users may download and/or print one copy of the publication from the University of Birmingham research portal for the purpose of private study or non-commercial research. •User may use extracts from the document in line with the concept of ‘fair dealing’ under the Copyright, Designs and Patents Act 1988 (?) •Users may not further distribute the material nor use it for the purposes of commercial gain. Where a licence is displayed above, please note the terms and conditions of the licence govern your use of this document. When citing, please reference the published version. Take down policy While the University of Birmingham exercises care and attention in making items available there are rare occasions when an item has been uploaded in error or has been deemed to be commercially or otherwise sensitive. -

ZNF750 Interacts with KLF4 and RCOR1, KDM1A, and CTBP1/2 Chromatin Regulators to Repress Epidermal Progenitor Genes and Induce Differentiation Genes
Downloaded from genesdev.cshlp.org on September 27, 2021 - Published by Cold Spring Harbor Laboratory Press ZNF750 interacts with KLF4 and RCOR1, KDM1A, and CTBP1/2 chromatin regulators to repress epidermal progenitor genes and induce differentiation genes Lisa D. Boxer,1,2 Brook Barajas,1 Shiying Tao,1 Jiajing Zhang,1 and Paul A. Khavari1,3 1Program in Epithelial Biology, Stanford University School of Medicine, Stanford, California 94305, USA; 2Department of Biology, Stanford University, Stanford, California 94305, USA; 3Veterans Affairs Palo Alto Healthcare System, Palo Alto, California 94304, USA ZNF750 controls epithelial homeostasis by inhibiting progenitor genes while inducing differentiation genes, a role underscored by pathogenic ZNF750 mutations in cancer and psoriasis. How ZNF750 accomplishes these dual gene regulatory impacts is unknown. Here, we characterized ZNF750 as a transcription factor that binds both the progenitor and differentiation genes that it controls at a CCNNAGGC DNA motif. ZNF750 interacts with the pluripotency transcription factor KLF4 and chromatin regulators RCOR1, KDM1A, and CTBP1/2 through conserved PLNLS sequences. ChIP-seq (chromatin immunoprecipitation [ChIP] followed by high-throughput sequencing) and gene depletion revealed that KLF4 colocalizes ~10 base pairs from ZNF750 at differentiation target genes to facilitate their activation but is unnecessary for ZNF750-mediated progenitor gene repression. In contrast, KDM1A colocalizes with ZNF750 at progenitor genes and facilitates their repression but is unnecessary for ZNF750-driven differentiation. ZNF750 thus controls differentiation in concert with RCOR1 and CTBP1/2 by acting with either KDM1A to repress progenitor genes or KLF4 to induce differentiation genes. [Keywords: stem cell; differentiation; ZNF750; KLF4; chromatin regulator] Supplemental material is available for this article. -

Y-Box Binding Protein 1 Is Up-Regulated in Proliferative Breast Cancer and Its Inhibition Deregulates the Cell Cycle
483-492.qxd 21/6/2010 12:04 ÌÌ ™ÂÏ›‰·483 INTERNATIONAL JOURNAL OF ONCOLOGY 37: 483-492, 2010 483 Y-box binding protein 1 is up-regulated in proliferative breast cancer and its inhibition deregulates the cell cycle YING-NAN YU1, GEORGE WAI-CHEONG YIP1, PUAY-HOON TAN1,2, AYE AYE THIKE2, KEN MATSUMOTO3, MASAFUMI TSUJIMOTO3 and BOON-HUAT BAY1 1Department of Anatomy, Yong Loo Lin School of Medicine, National University of Singapore; 2Department of Pathology, Singapore General Hospital, Singapore; 3Laboratory of Cellular Biochemistry, Institute of Physical and Chemical Research (RIKEN), Wako, Japan Received March 4, 2010; Accepted April 27, 2010 DOI: 10.3892/ijo_00000697 Abstract. The Y-box-binding protein 1 (YB-1), a member of Introduction the cold-shock domain RNA-and DNA-binding protein family, has pleiotropic functions such as regulation of the Breast cancer is the most frequent malignancy among women cell cycle. The aim of this study was to evaluate if YB-1 is a in Western countries. It is estimated that more than 1.1 million proliferative marker in breast cancer and elucidate potential new breast cancer cases are diagnosed worldwide per year (1). downstream targets involved in YB-1-mediated cell cycle Identification of clinically useful biomarkers is imperative regulation using RNA interference technology. YB-1 protein for optimal care and management of breast tumors (2). Several expression was evaluated in tissue microarrays of 131 breast proteins such as estrogen receptors, epidermal growth factor invasive ductal carcinomas by immunohistochemistry, while receptors, BRCA1 and progesterone receptors have been the YB-1 gene expression profile was evaluated in the T-47D, reported to be prognostic biomarkers in breast cancer (3,4). -

RBBP8 Antibody Cat
RBBP8 Antibody Cat. No.: 5761 Western blot analysis of RBBP8 in mouse spleen tissue lysate with RBBP8 antibody at (A) 1 and (B) 2 μg/mL. Immunohistochemistry of RBBP8 in rat spleen tissue with Immunofluorescence of RBBP8 in rat spleen tissue with RBBP8 antibody at 5 μg/mL. RBBP8 antibody at 20 μg/mL. Specifications HOST SPECIES: Rabbit SPECIES REACTIVITY: Human, Mouse, Rat RBBP8 antibody was raised against a 14 amino acid synthetic peptide near the center of human RBBP8. IMMUNOGEN: The immunogen is located within amino acids 430 - 480 of RBBP8. TESTED APPLICATIONS: ELISA, IF, IHC-P, WB September 27, 2021 1 https://www.prosci-inc.com/rbbp8-antibody-5761.html RBBP8 antibody can be used for detection of RBBP8 by Western blot at 1 - 2 μg/mL. Antibody can also be used for immunohistochemistry starting at 5 μg/mL. For immunofluorescence start at 20 μg/mL. APPLICATIONS: Antibody validated: Western Blot in mouse samples; Immunohistochemistry in rat samples and Immunofluorescence in rat samples. All other applications and species not yet tested. POSITIVE CONTROL: 1) Cat. No. 1406 - Mouse Spleen Tissue Lysate Properties PURIFICATION: RBBP8 Antibody is affinity chromatography purified via peptide column. CLONALITY: Polyclonal ISOTYPE: IgG CONJUGATE: Unconjugated PHYSICAL STATE: Liquid BUFFER: RBBP8 Antibody is supplied in PBS containing 0.02% sodium azide. CONCENTRATION: 1 mg/mL RBBP8 antibody can be stored at 4˚C for three months and -20˚C, stable for up to one STORAGE CONDITIONS: year. As with all antibodies care should be taken to avoid repeated freeze thaw cycles. Antibodies should not be exposed to prolonged high temperatures. -
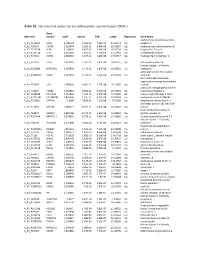
Table S1. the Statistical Metrics for Key Differentially Expressed Genes (Degs)
Table S1. The statistical metrics for key differentially expressed genes (DEGs) Gene Agilent Id Symbol logFC pValue FDR tvalue Regulation Gene Name oxidized low density lipoprotein A_24_P124624 OLR1 2.458429 1.19E-13 7.25E-10 24.04241 Up receptor 1 A_23_P90273 CHST8 2.622464 3.85E-12 6.96E-09 19.05867 Up carbohydrate sulfotransferase 8 A_23_P217528 KLF8 2.109007 4.85E-12 7.64E-09 18.76234 Up Kruppel like factor 8 A_23_P114740 CFH 2.651636 1.85E-11 1.79E-08 17.13652 Up complement factor H A_23_P34031 XAGE2 2.000935 2.04E-11 1.81E-08 17.02457 Up X antigen family member 2 A_23_P27332 TCF4 1.613097 2.32E-11 1.87E-08 16.87275 Up transcription factor 4 histone cluster 1 H1 family A_23_P250385 HIST1H1B 2.298658 2.47E-11 1.87E-08 16.80362 Up member b abnormal spindle microtubule A_33_P3288159 ASPM 2.162032 2.79E-11 2.01E-08 16.66292 Up assembly H19, imprinted maternally expressed transcript (non-protein A_24_P52697 H19 1.499364 4.09E-11 2.76E-08 16.23387 Up coding) potassium voltage-gated channel A_24_P31627 KCNB1 2.289689 6.65E-11 3.97E-08 15.70253 Up subfamily B member 1 A_23_P214168 COL12A1 2.155835 7.59E-11 4.15E-08 15.56005 Up collagen type XII alpha 1 chain A_33_P3271341 LOC388282 2.859496 7.61E-11 4.15E-08 15.55704 Up uncharacterized LOC388282 A_32_P150891 DIAPH3 2.2068 7.83E-11 4.22E-08 15.5268 Up diaphanous related formin 3 zinc finger protein 185 with LIM A_23_P11025 ZNF185 1.385721 8.74E-11 4.59E-08 15.41041 Up domain heat shock protein family B A_23_P96872 HSPB11 1.887166 8.94E-11 4.64E-08 15.38599 Up (small) member 11 A_23_P107454