CANPOLIN - Bee Course 2012
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Of Turkey Part II: the Genera Haetosmia, Osmia and Protosmia
Atatürk Üniv. Ziraat Fak. Derg., 44 (2): 121-143, 2013 Atatürk Univ., J. of the Agricultural Faculty, 44 (2): 121-143, 2013 ISSN : 1300-9036 Araştırma Makalesi/Research Article Distribution of the tribe Osmiini bees (Hymenoptera: Megachilidae) of Turkey Part II: the genera Haetosmia, Osmia and Protosmia Hikmet ÖZBEK Department of Plant Protection, Faculty of Agriculture, Atatürk Üniversity, Erzurum, Turkey ([email protected]) Geliş Tarihi : 23.03.2014 Kabul Tarihi : 17.12.2014 ABSTRACT: In the present study, research materials consisting of approximately 1200 bee samples, which have been collected from almost the entire country since the 1960s, but mainly from the eastern Anatolia region of Turkey, were evaluated, a total of 101 species and subspecies in three genera, Haetosmia (1), Osmia (87) and Protosmia (13) were recognized. Of these, Osmia (Helicosmia) labialis Pérez, 1879 is new for Turkey as well Asian Continenet. For the known species new distribution records were provided and the plant species visited were included. Majority of the Turkish species, about 70%, are associated with Europe, mainly Mediterranean basin. Therefore, many species are in Mediterranean and East-Mediterranean chorotypes. Where as about 30% are associated with Asia, most of them are in the Southwest-Asiatic chorotype, 13 species could be accepted as Anatolian endemics. Two taxa, Osmia (Hoplosmia) bidentata pallens (Tkalcu, 1979) and Protosmia (Protosmia) monstrosa (Pérez, 1895) occur in Asia and North Africa. Key words: Hymenoptera: Megachilidae, Osmiini, Haetosmia, Osmia, Protosmia, fauna, new record, chorotype, Turkey. Türkiye’de Osmiini tribüsüne giren arı türleri (Hymenoptera: Megachilidae) Bölüm II: Haetosmia, Osmia and Protosmia cinsleri Özet: Bu çalışmada; Türkiye’nin hemen her yöresinden, özellikle de Doğu Anadolu Bölgesi’nden 1960’lı yıllardan buyana toplanan 1200 civarındaki arı örneği değerlendirilmiş ve Haetosmia (1), Osmia (87 ) ve Protosmia (13) cinslerine ait 101 tür ve alttürün bulunduğu belirlenmiştir. -

Xhaie'ican%Mllsllm
XhAie'ican1ox4tate%Mllsllm PUBLISHED BY THE AMERICAN MUSEUM OF NATURAL HISTORY CENTRAL PARK WEST AT 79TH STREET, NEW YORK 24, N.Y. NUMBER 2 244 MAY I9, I 966 The Larvae of the Anthophoridae (Hymenoptera, Apoidea) Part 2. The Nomadinae BY JEROME G. ROZEN, JR.1 The present paper is the second of a series that treats the phylogeny and taxonomy of the larvae belonging to the bee family Anthophoridae. The first (Rozen, 1965a) dealt with the pollen-collecting tribes Eucerini and Centridini of the Anthophorinae. The present study encompasses the following tribes, all of which consist solely of cuckoo bees: Protepeolini, Epeolini, Nomadini, Ammobatini, Holcopasitini, Biastini, and Neolarrini. For reasons presented below, these tribes are believed to represent a monophyletic group, and consequently all are placed in the Nomadinae. It seems likely that the cleptoparasitic tribes Caenoprosopini, Ammoba- toidini, Townsendiellini, Epeoloidini, and Osirini are also members of the subfamily, although their larvae have not as yet been collected. Although the interrelationships of the numerous taxa within the Nomadinae need to be re-evaluated, the tribal concepts used by Michener (1944) are employed here. Adjustments in the classifications will certainly have to be made in the future, however, for Michener (1954) has already indicated, for example, that characters of the adults in the Osirini, the Epeolini, and the Nomadini intergrade. The affinities of the Nomadinae with the other subfamilies of the Antho- phoridae will be discussed in the last paper of the series. Because of char- 1 Curator, Department of Entomology, the American Museum of Natural History. 2 AMERICAN MUSEUM NOVITATES NO. -

Taxon Order Family Scientific Name Common Name Non-Native No. of Individuals/Abundance Notes Bees Hymenoptera Andrenidae Calliop
Taxon Order Family Scientific Name Common Name Non-native No. of individuals/abundance Notes Bees Hymenoptera Andrenidae Calliopsis andreniformis Mining bee 5 Bees Hymenoptera Apidae Apis millifera European honey bee X 20 Bees Hymenoptera Apidae Bombus griseocollis Brown belted bumble bee 1 Bees Hymenoptera Apidae Bombus impatiens Common eastern bumble bee 12 Bees Hymenoptera Apidae Ceratina calcarata Small carpenter bee 9 Bees Hymenoptera Apidae Ceratina mikmaqi Small carpenter bee 4 Bees Hymenoptera Apidae Ceratina strenua Small carpenter bee 10 Bees Hymenoptera Apidae Melissodes druriella Small carpenter bee 6 Bees Hymenoptera Apidae Xylocopa virginica Eastern carpenter bee 1 Bees Hymenoptera Colletidae Hylaeus affinis masked face bee 6 Bees Hymenoptera Colletidae Hylaeus mesillae masked face bee 3 Bees Hymenoptera Colletidae Hylaeus modestus masked face bee 2 Bees Hymenoptera Halictidae Agapostemon virescens Sweat bee 7 Bees Hymenoptera Halictidae Augochlora pura Sweat bee 1 Bees Hymenoptera Halictidae Augochloropsis metallica metallica Sweat bee 2 Bees Hymenoptera Halictidae Halictus confusus Sweat bee 7 Bees Hymenoptera Halictidae Halictus ligatus Sweat bee 2 Bees Hymenoptera Halictidae Lasioglossum anomalum Sweat bee 1 Bees Hymenoptera Halictidae Lasioglossum ellissiae Sweat bee 1 Bees Hymenoptera Halictidae Lasioglossum laevissimum Sweat bee 1 Bees Hymenoptera Halictidae Lasioglossum platyparium Cuckoo sweat bee 1 Bees Hymenoptera Halictidae Lasioglossum versatum Sweat bee 6 Beetles Coleoptera Carabidae Agonum sp. A ground beetle -

Pollination of Cultivated Plants in the Tropics 111 Rrun.-Co Lcfcnow!Cdgmencle
ISSN 1010-1365 0 AGRICULTURAL Pollination of SERVICES cultivated plants BUL IN in the tropics 118 Food and Agriculture Organization of the United Nations FAO 6-lina AGRICULTUTZ4U. ionof SERNES cultivated plans in tetropics Edited by David W. Roubik Smithsonian Tropical Research Institute Balboa, Panama Food and Agriculture Organization of the United Nations F'Ø Rome, 1995 The designations employed and the presentation of material in this publication do not imply the expression of any opinion whatsoever on the part of the Food and Agriculture Organization of the United Nations concerning the legal status of any country, territory, city or area or of its authorities, or concerning the delimitation of its frontiers or boundaries. M-11 ISBN 92-5-103659-4 All rights reserved. No part of this publication may be reproduced, stored in a retrieval system, or transmitted in any form or by any means, electronic, mechanical, photocopying or otherwise, without the prior permission of the copyright owner. Applications for such permission, with a statement of the purpose and extent of the reproduction, should be addressed to the Director, Publications Division, Food and Agriculture Organization of the United Nations, Viale delle Terme di Caracalla, 00100 Rome, Italy. FAO 1995 PlELi. uion are ted PlauAr David W. Roubilli (edita Footli-anal ISgt-iieulture Organization of the Untled Nations Contributors Marco Accorti Makhdzir Mardan Istituto Sperimentale per la Zoologia Agraria Universiti Pertanian Malaysia Cascine del Ricci° Malaysian Bee Research Development Team 50125 Firenze, Italy 43400 Serdang, Selangor, Malaysia Stephen L. Buchmann John K. S. Mbaya United States Department of Agriculture National Beekeeping Station Carl Hayden Bee Research Center P. -

Wedge-Shaped Beetles (Suggested Common Name) Ripiphorus Spp. (Insecta: Coleoptera: Ripiphoridae)1 David Owens, Ashley N
EENY613 Wedge-Shaped Beetles (suggested common name) Ripiphorus spp. (Insecta: Coleoptera: Ripiphoridae)1 David Owens, Ashley N. Mortensen, Jeanette Klopchin, William Kern, and Jamie D. Ellis2 Introduction Ripiphoridae are a family of unusual parasitic beetles that are thought to be related to tumbling flower beetles (Coleoptera: Mordellidae) and blister beetles (Coleoptera: Meloidae). There is disagreement over the spelling of the family (Ripiphoridae) and genus (Ripiphorus) names. Here we use the original spelling that starts with only the letter Figure 1. Adult specimens of the two genera of Ripiphoridae. A) “R”; however, an initial “Rh” has also been used in the Macrosiagon Hentz, and B) Ripiphorus Bosc. scientific community (Rhipiphoridae and Rhipiphorus). Credits: Allen M. Boatman There are an estimated 35 nearctic species of Ripiphorus, Generally, the biology of the family Ripiphoridae is poorly two of which have been collected in Florida: Ripiphorus known. Ripiphorids parasitize bees and wasps (Hymenop- schwarzi LeConte (Figure 2A) and Ripiphorus fasciatus Say tera), roaches (Blattodea), and wood-boring beetles (Co- (Figure 2B). Due to limited information for both of these leoptera). However, the specific hosts for many ripiphorid species, the information presented below is characteristic species are unknown. Furthermore, only one sex (either of the genus Ripiphorus. Information specific to Ripiphorus male or female) has been described for several species, and fasciatus and Ripiphorus schwarzi is presented where the males and females of some species look different. detailed information is available. Two genera of Ripiphoridae infest hymenopteran (bee and wasp) nests: Macrosiagon Hentz (Figure 1A) and Ripiphorus Bosc (formerly Myodites Latreille) (Figure 1B). Species of Macrosiagon are parasites of a variety of hymenopteran families including: Halictidae, Vespidae, Tiphiidae, Apidae, Pompilidae, Crabronidae, and Sphecidae. -

Apoidea: Andrenidae: Panurginae)
Contemporary distributions of Panurginus species and subspecies in Europe (Apoidea: Andrenidae: Panurginae) Sébastien Patiny Abstract The largest number of Old World Panurginus Nylander, 1848 species is distributed in the West- Palaearctic. The genus is absent in Africa and rather rare in the East-Palaearctic. Warncke (1972, 1987), who is the main author treating Palaearctic Panurginae in the last decades, subdivided Panurginus into a small number of species, including two principal taxa admitting for each a very large number of subspecies: Panurginus brullei (Lepeletier, 1841) and Panurginus montanus Giraud, 1861. Following recent works, the two latter are in fact complexes of closely related species. In the West-Palaearctic context, distributions of certain species implied in these complexes appear as very singular, distinct of mostly all other Panurginae ranges and highly interesting from a fundamental point of view. Based on a cartographic approach, the causes which influence (or have conditioned, in the past) the observed ranges of these species are discussed. Key words: Panurginae, biogeography, West-Palaearctic, speciation, glaciation. Introduction 1841) sensu lato are studied in the present paper. In the Old World Panurginae fauna, Panurginus The numerous particularities of their distribu- Nylander, 1848 is one of the only two genera tions are characterized and discussed here. The (with Melitturga Latreille, 1809) which are limits of these distributions were proposed and distributed in the entire Palaearctic region. The related to the contemporary and past develop- genera Camptopoeum Spinola, 1843 and Panur- ments which could have caused these ranges. gus Panzer, 1806 are also represented in the East- Palaearctic (in northern Thailand), but too few Catalogue of the old world species data are available for these genera to make them of Panurginus the subject of particular considerations. -
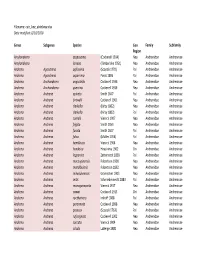
Anthophila List
Filename: cuic_bee_database. -

Redalyc.CLEPTOPARASITE BEES, with EMPHASIS on THE
Acta Biológica Colombiana ISSN: 0120-548X [email protected] Universidad Nacional de Colombia Sede Bogotá Colombia ALVES-DOS-SANTOS, ISABEL CLEPTOPARASITE BEES, WITH EMPHASIS ON THE OILBEES HOSTS Acta Biológica Colombiana, vol. 14, núm. 2, 2009, pp. 107-113 Universidad Nacional de Colombia Sede Bogotá Bogotá, Colombia Available in: http://www.redalyc.org/articulo.oa?id=319027883009 How to cite Complete issue Scientific Information System More information about this article Network of Scientific Journals from Latin America, the Caribbean, Spain and Portugal Journal's homepage in redalyc.org Non-profit academic project, developed under the open access initiative Acta biol. Colomb., Vol. 14 No. 2, 2009 107 - 114 CLEPTOPARASITE BEES, WITH EMPHASIS ON THE OILBEES HOSTS Abejas cleptoparásitas, con énfasis en las abejas hospederas coletoras de aceite ISABEL ALVES-DOS-SANTOS1, Ph. D. 1Departamento de Ecologia, IBUSP. Universidade de São Paulo, Rua do Matão 321, trav 14. São Paulo 05508-900 Brazil. [email protected] Presentado 1 de noviembre de 2008, aceptado 1 de febrero de 2009, correcciones 7 de julio de 2009. ABSTRACT Cleptoparasite bees lay their eggs inside nests constructed by other bee species and the larvae feed on pollen provided by the host, in this case, solitary bees. The cleptoparasite (adult and larvae) show many morphological and behavior adaptations to this life style. In this paper I present some data on the cleptoparasite bees whose hosts are bees specialized to collect floral oil. Key words: solitary bee, interspecific interaction, parasitic strategies, hospicidal larvae. RESUMEN Las abejas Cleptoparásitas depositan sus huevos en nidos construídos por otras especies de abejas y las larvas se alimentan del polen que proveen las hospederas, en este caso, abejas solitarias. -

Evolution of the Suctorial Proboscis in Pollen Wasps (Masarinae, Vespidae)
Arthropod Structure & Development 31 (2002) 103–120 www.elsevier.com/locate/asd Evolution of the suctorial proboscis in pollen wasps (Masarinae, Vespidae) Harald W. Krenna,*, Volker Maussb, John Planta aInstitut fu¨r Zoologie, Universita¨t Wien, Althanstraße 14, A-1090, Vienna, Austria bStaatliches Museum fu¨r Naturkunde, Abt. Entomologie, Rosenstein 1, D-70191 Stuttgart, Germany Received 7 May 2002; accepted 17 July 2002 Abstract The morphology and functional anatomy of the mouthparts of pollen wasps (Masarinae, Hymenoptera) are examined by dissection, light microscopy and scanning electron microscopy, supplemented by field observations of flower visiting behavior. This paper focuses on the evolution of the long suctorial proboscis in pollen wasps, which is formed by the glossa, in context with nectar feeding from narrow and deep corolla of flowers. Morphological innovations are described for flower visiting insects, in particular for Masarinae, that are crucial for the production of a long proboscis such as the formation of a closed, air-tight food tube, specializations in the apical intake region, modification of the basal articulation of the glossa, and novel means of retraction, extension and storage of the elongated parts. A cladistic analysis provides a framework to reconstruct the general pathways of proboscis evolution in pollen wasps. The elongation of the proboscis in context with nectar and pollen feeding is discussed for aculeate Hymenoptera. q 2002 Elsevier Science Ltd. All rights reserved. Keywords: Mouthparts; Flower visiting; Functional anatomy; Morphological innovation; Evolution; Cladistics; Hymenoptera 1. Introduction Some have very long proboscides; however, in contrast to bees, the proboscis is formed only by the glossa and, in Evolution of elongate suctorial mouthparts have some species, it is looped back into the prementum when in occurred separately in several lineages of Hymenoptera in repose (Bradley, 1922; Schremmer, 1961; Richards, 1962; association with uptake of floral nectar. -

Hymenoptera: Apidae), with Distributional Modeling of Adventive Euglossines
Comparative Genital Morphology, Phylogeny, and Classification of the Orchid Bee Genus Euglossa Latreille (Hymenoptera: Apidae), with Distributional Modeling of Adventive Euglossines BY ©2010 Ismael Alejandro Hinojosa Díaz Submitted to the graduate degree program in Ecology and Evolutionary Biology and the Graduate Faculty of the University of Kansas in partial fulfillment of the requirements for the degree of Doctor of Philosophy. Chairperson Michael S. Engel Charles D. Michener Edward O. Wiley Kirsten Jensen J. Christopher Brown Date Defended: November 10, 2010 The Dissertation Committee for Ismael Alejandro Hinojosa Díaz certifies that this is the approved version of the following dissertation: Comparative Genital Morphology, Phylogeny, and Classification of the Orchid Bee Genus Euglossa Latreille (Hymenoptera: Apidae), with Distributional Modeling of Adventive Euglossines Chairperson Michael S. Engel Date approved: November 22, 2010 ii ABSTRACT Orchid bees (tribe Euglossini) are conspicuous members of the corbiculate bees owing to their metallic coloration, long labiomaxillary complex, and the fragrance-collecting behavior of the males, more prominently (but not restricted) from orchid flowers (hence the name of the group). They are the only corbiculate tribe that is exclusively Neotropical and without eusocial members. Of the five genera in the tribe, Euglossa Latreille is the most diverse with around 120 species. Taxonomic work on this genus has been linked historically to the noteworthy secondary sexual characters of the males, which combined with the other notable external features, served as a basis for the subgeneric classification commonly employed. The six subgenera Dasystilbe Dressler, Euglossa sensu stricto, Euglossella Moure, Glossura Cockerell, Glossurella Dressler and Glossuropoda Moure, although functional for the most part, showed some intergradations (especially the last three), and no phylogenetic evaluation of their validity has been produced. -

Efectos De La Fragmentación Del Hábitat Sobre Himenópteros Antófilos (Insecta) En El Bosque Chaqueño Serrano
Universidad Nacional de Córdoba Facultad de Ciencias Exactas Físicas y Naturales Doctorado en Ciencias Biológicas Manuscrito de Tesis para optar al título de Dra. en Ciencias Biológicas Efectos de la fragmentación del hábitat sobre himenópteros antófilos (Insecta) en el Bosque Chaqueño Serrano Doctorando: Bióloga Mariana Laura Musicante Directora: Dra. Adriana Salvo Co-Director: Dr. Leonardo Galetto Centro de Investigaciones Entomológicas de Córdoba (CIEC) Instituto Multidisciplinario de Biología Vegetal (IMBIV-CONICET) Córdoba, Argentina 2013 Comisión Asesora Dr. Marcelo Aizen Laboratorio Ecotono-Centro Regional Universitario Bariloche (CRUB), Universidad Nacional del Comahue e Instituto de Investigaciones en Biodiversidad y Medioambiente (INIBIOMA), San Carlos de Bariloche. Departamento de Botánica, Museo Argentino de Ciencias Naturales, Buenos Aires. Dr. Marcelo Cabido Instituto Multidisciplinario de Biología Vegetal-CONICT. Universidad Nacional de Córdoba. Dra. Adriana Salvo Centro de Investigaciones Entomológicas de Córdoba. Instituto Multidisciplinario de Biología Vegetal-CONICT Universidad Nacional de Córdoba. Defensa Oral y Pública Lugar y fecha: Calificación: Tribunal ______________________________ _____________________________________ Firma Aclaración ______________________________ _____________________________________ Firma Aclaración ______________________________ ____________________________________ Firma Aclaración A esos pequeños seres que zumbaban ayer y a los que todavía zumban hoy Efectos de la fragmentación del hábitat -

Identification of 37 Microsatellite Loci for Anthophora Plumipes (Hymenoptera: Apidae) Using Next Generation Sequencing and Their Utility in Related Species
Eur. J. Entomol. 109: 155–160, 2012 http://www.eje.cz/scripts/viewabstract.php?abstract=1692 ISSN 1210-5759 (print), 1802-8829 (online) Identification of 37 microsatellite loci for Anthophora plumipes (Hymenoptera: Apidae) using next generation sequencing and their utility in related species KATEěINA ýERNÁ and JAKUB STRAKA Department of Zoology, Faculty of Science, Charles University in Prague, Viniþná 7, 128 43 Praha 2, Czech Republic; e-mails: [email protected]; [email protected] Key words. Hymenoptera, Apidae, microsatellite development, Anthophora plumipes, 454 sequencing, Anthophorini Abstract. Novel microsatellite markers for the solitary bee, Anthophora plumipes, were identified and characterised using 454 GS-FLX Titanium pyrosequencing technology. Thirty seven loci were tested using fluorescently labelled primers on a sample of 20 females from Prague. The number of alleles ranged from 1 to 10 (with a mean of 4 alleles per locus), resulting in an observed hetero- zygosity ranging from 0.05 to 0.9 and an expected heterozygosity from 0.097 to 0.887. None of the loci showed a significant devia- tion from the Hardy-Weinberg equilibrium and only two loci showed the significant presence of null alleles. No linkage between loci was detected. We further provide information on a single multiplex PCR consisting of 11 of the most polymorphic loci. This multi- plex approach provides an effective analytical tool for analysing genetic structure and carrying out parental analyses on Anthophora populations. Most of the 37 loci tested also showed robust amplification in five other Anthophora species (A. aestivalis, A. crinipes, A. plagiata, A. pubescens and A. quadrimaculata).