Cholinesterase Inhibitors from Plants: Possible Treatment Strategy for Neurological Disorders – a Review
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Protein Complex Formation by Acetylcholinesterase and the Neurotoxin Fasciculin-2 Appears to Involve an Induced-Fit Mechanism
Protein complex formation by acetylcholinesterase and the neurotoxin fasciculin-2 appears to involve an induced-fit mechanism Jennifer M. Bui†‡ and J. Andrew McCammon†§ †Department of Chemistry and Biochemistry, Howard Hughes Medical Institute, and §Department of Pharmacology, University of California at San Diego, 9500 Gilman Drive, La Jolla, CA 92093-0365 Edited by Jose N. Onuchic, University of California at San Diego, La Jolla, CA, and approved August 22, 2006 (received for review June 27, 2006) Specific, rapid association of protein complexes is essential for all forms of cellular existence. The initial association of two molecules in diffusion-controlled reactions is often influenced by the elec- trostatic potential. Yet, the detailed binding mechanisms of pro- teins highly depend on the particular system. A complete protein complex formation pathway has been delineated by using struc- tural information sampled over the course of the transformation reaction. The pathway begins at an encounter complex that is formed by one of the apo forms of neurotoxin fasciculin-2 (FAS2) and its high-affinity binding protein, acetylcholinesterase (AChE), followed by rapid conformational rearrangements into an inter- mediate complex that subsequently converts to the final complex as observed in crystal structures. Formation of the intermediate complex has also been independently captured in a separate 20-ns Fig. 1. Thermodynamic cycle for AB* complex formation reactions. A and B BIOPHYSICS molecular dynamics simulation of the encounter complex. Confor- molecules can be considered as any pair of interacting molecules. mational transitions between the apo and liganded states of FAS2 in the presence and absence of AChE are described in terms of their relative free energy profiles that link these two states. -

Antioxidants and Second Messengers of Free Radicals
antioxidants Antioxidants and Second Messengers of Free Radicals Edited by Neven Zarkovic Printed Edition of the Special Issue Published in Antioxidants www.mdpi.com/journal/antioxidants Antioxidants and Second Messengers of Free Radicals Antioxidants and Second Messengers of Free Radicals Special Issue Editor Neven Zarkovic MDPI • Basel • Beijing • Wuhan • Barcelona • Belgrade Special Issue Editor Neven Zarkovic Rudjer Boskovic Institute Croatia Editorial Office MDPI St. Alban-Anlage 66 4052 Basel, Switzerland This is a reprint of articles from the Special Issue published online in the open access journal Antioxidants (ISSN 2076-3921) from 2018 (available at: https://www.mdpi.com/journal/ antioxidants/special issues/second messengers free radicals) For citation purposes, cite each article independently as indicated on the article page online and as indicated below: LastName, A.A.; LastName, B.B.; LastName, C.C. Article Title. Journal Name Year, Article Number, Page Range. ISBN 978-3-03897-533-5 (Pbk) ISBN 978-3-03897-534-2 (PDF) c 2019 by the authors. Articles in this book are Open Access and distributed under the Creative Commons Attribution (CC BY) license, which allows users to download, copy and build upon published articles, as long as the author and publisher are properly credited, which ensures maximum dissemination and a wider impact of our publications. The book as a whole is distributed by MDPI under the terms and conditions of the Creative Commons license CC BY-NC-ND. Contents About the Special Issue Editor ...................................... vii Preface to ”Antioxidants and Second Messengers of Free Radicals” ................ ix Neven Zarkovic Antioxidants and Second Messengers of Free Radicals Reprinted from: Antioxidants 2018, 7, 158, doi:10.3390/antiox7110158 ............... -

Β-Phenylethylamines and the Isoquinoline Alkaloids
-Phenylethylamines and the isoquinoline alkaloids Kenneth W. Bentley Marrview, Tillybirloch, Midmar, Aberdeenshire, UK AB51 7PS Received (in Cambridge, UK) 28th November 2000 First published as an Advance Article on the web 7th February 2001 Covering: July 1999 to June 2000. Previous review: Nat. Prod. Rep., 2000, 17, 247. 1 Introduction 2 -Phenylethylamines 3 Isoquinolines 4 Naphthylisoquinolines 5 Benzylisoquinolines 6 Bisbenzylisoquinolines 7 Pavines and isopavines 8 Berberines and tetrahydoberberines 9 Protopines 10 Phthalide-isoquinolines 11 Other modified berberines 12 Emetine and related alkaloids 13 Benzophenanthridines 14 Aporphinoid alkaloids 14.1 Proaporphines 14.2 Aporphines 14.3 Aporphine–benzylisoquinoline dimers 14.4 Phenanthrenes 14.5 Oxoaporphines 14.6 Dioxoaporphines 14.7 Aristolochic acids and aristolactams 14.8 Oxoisoaporphines 15 Alkaloids of the morphine group 16 Colchicine and related alkaloids 17 Erythrina alkaloids 17.1 Erythrinanes 17.2 Cephalotaxine and related alkaloids 18 Other isoquinolines 19 References 1 Introduction Reviews of the occurrence of isoquinoline alkaloids in some plant species 1,2 and of recent developments in the chemistry and synthesis of alkaloids of these groups 3–6 have been published. 2 -Phenylethylamines β-Phenylethylamine, tyramine, N-methyltyramine, hordenine, mescaline, N-methylmescaline and N,N-dimethylmescaline 1, which is reported as an alkaloid for the first time, have been isolated from an unspecified species of Turbinocarpus 7 and N-trans-feruloyltyramine has been isolated from Cananga odorata.8 The N-oxides of the known alkaloid culantraramine 2 and the unknown culantraraminol 3, together with the related avicennamine 4 have been isolated as new alkaloids from Zanthoxylum avicennae.9 Three novel amides of dehydrotyr- leucine and proline respectively. -

Comparison of the Binding of Reversible Inhibitors to Human Butyrylcholinesterase and Acetylcholinesterase: a Crystallographic, Kinetic and Calorimetric Study
Article Comparison of the Binding of Reversible Inhibitors to Human Butyrylcholinesterase and Acetylcholinesterase: A Crystallographic, Kinetic and Calorimetric Study Terrone L. Rosenberry 1, Xavier Brazzolotto 2, Ian R. Macdonald 3, Marielle Wandhammer 2, Marie Trovaslet-Leroy 2,†, Sultan Darvesh 4,5,6 and Florian Nachon 2,* 1 Departments of Neuroscience and Pharmacology, Mayo Clinic College of Medicine, Jacksonville, FL 32224, USA; [email protected] 2 Département de Toxicologie et Risques Chimiques, Institut de Recherche Biomédicale des Armées, 91220 Brétigny-sur-Orge, France; [email protected] (X.B.); [email protected] (M.W.); [email protected] (M.T.-L.) 3 Department of Diagnostic Radiology, Dalhousie University, Halifax, NS B3H 4R2, Canada; [email protected] 4 Department of Medical Neuroscience, Dalhousie University, Halifax, NS B3H 4R2, Canada; [email protected] 5 Department of Chemistry, Mount Saint Vincent University, Halifax, NS B3M 2J6, Canada 6 Department of Medicine (Neurology and Geriatric Medicine), Dalhousie University, Halifax, NS B3H 4R2, Canada * Correspondence: [email protected]; Tel.: +33-178-65-1877 † Deceased October 2016. Received: 26 October 2017; Accepted: 27 November 2017; Published: 29 November 2017 Abstract: Acetylcholinesterase (AChE) and butyrylcholinesterase (BChE) hydrolyze the neurotransmitter acetylcholine and, thereby, function as coregulators of cholinergic neurotransmission. Although closely related, these enzymes display very different substrate specificities that only partially overlap. This disparity is largely due to differences in the number of aromatic residues lining the active site gorge, which leads to large differences in the shape of the gorge and potentially to distinct interactions with an individual ligand. Considerable structural information is available for the binding of a wide diversity of ligands to AChE. -

Anti-Cholinergic Alkaloids As Potential Therapeutic Agents for Alzheimer's Disease
Indian Journal of Biochemistry & Biophysics Vol. 50, April 2013, pp. 120-125 Anti-cholinergic alkaloids as potential therapeutic agents for Alzheimer’s disease: An in silico approach Huma Naaz, Swati Singh, Veda P Pandey, Priyanka Singh and Upendra N Dwivedi* Bioinformatics Infrastructure Facility, Center of Excellence in Bioinformatics, Department of Biochemistry, University of Lucknow, Lucknow 226 007, India Received 10 September 2012; revised 25 January 2013 Alzheimer’s disease (AD), a progressive neurodegenerative disorder with many cognitive and neuropsychiatric symptoms is biochemically characterized by a significant decrease in the brain neurotransmitter acetylcholine (ACh). Plant-derived metabolites, including alkaloids have been reported to possess neuroprotective properties and are considered to be safe, thus have potential for developing effective therapeutic molecules for neurological disorders, such as AD. Therefore, in the present study, thirteen plant-derived alkaloids, namely pleiocarpine, kopsinine, pleiocarpamine (from Pleiocarpa mutica, family: Annonaceae), oliveroline, noroliveroline, liridonine, isooncodine, polyfothine, darienine (from Polyalthia longifolia, family: Apocynaceae) and eburnamine, eburnamonine, eburnamenine and geissoschizol (from Hunteria zeylanica, family: Apocynaceae) were analyzed for their anti-cholinergic action through docking with acetylcholinesterase (AChE) as target. Among the alkaloids, pleiocarpine showed promising anti-cholinergic potential, while its amino derivative showed about six-fold -

Relief of Hypersensitivity After Nerve Injury from Systemic Donepezil
Saranya devi Relief of Hypersensitivity after Nerve Injury from ALN Systemic Donepezil Involves Spinal Cholinergic and γ-Aminobutyric Acid Mechanisms Spinal ACh and GABA Mechanisms for Donepezil Analgesia Masafumi Kimura, M.D.,* Ken-ichiro Hayashida, D.V.M., Ph.D.,† James C. Eisenach, M.D.,‡ KIMURA ET AL. Shigeru Saito M.D.,§ Hideaki Obata, M.D.‖ XX ABSTRACT What We Already Know about This Topic • Donepezil increases central nervous system acetylcholine lev- Downloaded from http://pubs.asahq.org/anesthesiology/article-pdf/118/1/173/259568/0000542-201301000-00030.pdf by guest on 28 September 2021 10.1097/ALN.0b013e318277a81c Background: Evoking spinal release of acetylcholine (ACh) els and is used for treatment of dementia produces antinociception in normal animals and reduces • Increases in spinal acetylcholine levels have been implicated in the relief of neuropathic pain hypersensitivity after nerve injury, and some studies suggest January that ACh-mediated analgesia relies on γ-aminobutyric acid (GABA)-ergic signaling in the spinal cord. In this study, the authors tested the spinal mechanisms underlying the anti- What This Article Tells Us That Is New 118 hypersensitivity effects of donepezil, a central nervous sys- • Donepezil increased spinal acetylcholine levels and γ- tem–penetrating cholinesterase inhibitor, in a rat model of aminobutyric acid levels to reduce nociceptive responses after nerve injury and represents a potential therapeutic pathway to neuropathic pain. reduce pain after nerve injury Methods: Male Sprague-Dawley rats were anesthetized, and L5 spinal nerve ligation was performed unilaterally. With- A antagonist; and CGP 35348 (30 μg), a γ-aminobutyric drawal threshold to a paw pressure test was measured before acid receptor type B antagonist. -

Tor Amendment Tagoor Laboratories Private Limited
ToR Amendment Tagoor Laboratories Private Limited ANNEXURE – I Details of Products as per Granted ToR and Amendment Required Production Capacity S. Ton /Month Therapeutic Product Name No As per Amendment category granted ToR Required 1 Abacavir sulfate 2.0 2.0 Anti HIV 2 Amitriptyline 5.0 10.0 Antidepressant 3 Atrovastatin Calcium 5.0 5.0 Hypercholesterolemia 4 Bupropion 5.0 5.0 Anti depressant 5 carisoprodol 2.0 2.0 Muscle Relaxant 6 Clopidogrelbisulfate 5.0 5.0 Antithrombotic 7 Cyclobenzaprine HCl 5.0 5.0 Muscle relaxant 8 Cyproheptadine HCl 5.0 10.0 Anti allergic 9 Desloratadine 5.0 5.0 Antihistamine 10 Domperidone 20.0 30.0 Anti emetic 11 Domperidone maleate 2.0 2.0 Anti emetic 12 Donepezil HCl 1.0 1.0 Alzheimer’s disease 13 Ebastine 5.0 5.0 Anti allergic 14 Esomeprazole Sodium 3.0 3.0 Anti ulcerative Esomeprazole Magnesium 3.0 3.0 Anti ulcerative 15 trihydrate 16 Fexofenadine Hydrochloride 6.0 15.0 Anti histamine 17 Haloperidol 2.0 2.0 Antipsychotic 18 Itopride Hydrochloride 2.0 2.0 Antispasmodics 19 Itraconazole 10.0 10.0 Antifungal 20 Ketrolac Tromethane 2.0 2.0 Anti Inflammatory 21 Lansoprazole 10.0 10.0 Ant ulcerative 22 Loperamide Hydrochloride 10.0 10.0 Anti diarrhea agent 23 Losartan Potassium 6.0 15.0 Anti Hypertensive 24 Nebivolol HCl 2.0 2.0 Anti Hypertensive 25 Nortriptyline HCl 2.0 2.0 Anti depressant 26 Omeprazole 40.0 40.0 Anti ulcerative 27 Omeprazole Sodium 2.0 2.0 Ant ulcerative Omeprazole Magnesium 2.0 2.0 Ant ulcerative 28 Dihydrate 29 Oxatomide 1.0 1.0 Antihistamine Pantoprazole Sodium Sesqui 10.0 20.0 Ant ulcerative 30 Hydrate 31 Pimozide 2.0 2.0 Antipsychotic 32 Pregabalin 2.0 2.0 Epileptic 33 Quetiapine Hemifumarate 2.0 2.0 Antipsychotic Prepared By Rightsource Industrial Solutions Pvt. -

Title 16. Crimes and Offenses Chapter 13. Controlled Substances Article 1
TITLE 16. CRIMES AND OFFENSES CHAPTER 13. CONTROLLED SUBSTANCES ARTICLE 1. GENERAL PROVISIONS § 16-13-1. Drug related objects (a) As used in this Code section, the term: (1) "Controlled substance" shall have the same meaning as defined in Article 2 of this chapter, relating to controlled substances. For the purposes of this Code section, the term "controlled substance" shall include marijuana as defined by paragraph (16) of Code Section 16-13-21. (2) "Dangerous drug" shall have the same meaning as defined in Article 3 of this chapter, relating to dangerous drugs. (3) "Drug related object" means any machine, instrument, tool, equipment, contrivance, or device which an average person would reasonably conclude is intended to be used for one or more of the following purposes: (A) To introduce into the human body any dangerous drug or controlled substance under circumstances in violation of the laws of this state; (B) To enhance the effect on the human body of any dangerous drug or controlled substance under circumstances in violation of the laws of this state; (C) To conceal any quantity of any dangerous drug or controlled substance under circumstances in violation of the laws of this state; or (D) To test the strength, effectiveness, or purity of any dangerous drug or controlled substance under circumstances in violation of the laws of this state. (4) "Knowingly" means having general knowledge that a machine, instrument, tool, item of equipment, contrivance, or device is a drug related object or having reasonable grounds to believe that any such object is or may, to an average person, appear to be a drug related object. -

An in Silico Study of the Ligand Binding to Human Cytochrome P450 2D6
AN IN SILICO STUDY OF THE LIGAND BINDING TO HUMAN CYTOCHROME P450 2D6 Sui-Lin Mo (Doctor of Philosophy) Discipline of Chinese Medicine School of Health Sciences RMIT University, Victoria, Australia January 2011 i Declaration I hereby declare that this submission is my own work and to the best of my knowledge it contains no materials previously published or written by another person, or substantial proportions of material which have been accepted for the award of any other degree or diploma at RMIT university or any other educational institution, except where due acknowledgment is made in the thesis. Any contribution made to the research by others, with whom I have worked at RMIT university or elsewhere, is explicitly acknowledged in the thesis. I also declare that the intellectual content of this thesis is the product of my own work, except to the extent that assistance from others in the project‘s design and conception or in style, presentation and linguistic expression is acknowledged. PhD Candidate: Sui-Lin Mo Date: January 2011 ii Acknowledgements I would like to take this opportunity to express my gratitude to my supervisor, Professor Shu-Feng Zhou, for his excellent supervision. I thank him for his kindness, encouragement, patience, enthusiasm, ideas, and comments and for the opportunity that he has given me. I thank my co-supervisor, A/Prof. Chun-Guang Li, for his valuable support, suggestions, comments, which have contributed towards the success of this thesis. I express my great respect to Prof. Min Huang, Dean of School of Pharmaceutical Sciences at Sun Yat-sen University in P.R.China, for his valuable support. -

EVALUATION of ACETYLCHOLINESTERASE INHIBITORY ACTIVITY and MODULATORY EFFECT on NICOTINIC ACETYLCHOLINE RECEPTORS of ALKALOIDS ISOLATED from Rhodolirium Andicola
UNIVERSIDAD DE LA FRONTERA Facultad de Ingeniería y Ciencias Doctorado en Ciencias de Recursos Naturales EVALUATION OF ACETYLCHOLINESTERASE INHIBITORY ACTIVITY AND MODULATORY EFFECT ON NICOTINIC ACETYLCHOLINE RECEPTORS OF ALKALOIDS ISOLATED FROM Rhodolirium andicola DOCTORAL THESIS IN FULFILLMENT OF THE REQUERIMENTS FOR THE DEGREE DOCTOR OF SCIENCES IN NATURAL RESOURCES FELIPE EDUARDO MORAGA NICOLÁS TEMUCO-CHILE 2018 Evaluation of acetylcholinesterase inhibitory activity and modulatory effect on nicotinic acetylcholine receptors of alkaloids isolated from Rhodolirium andicola Esta tesis fue realizada bajo la supervisión del director de Tesis Dra. ANA MUTIS TEJOS, y bajo la co-tutela del Dr. EMILIO HORMAZÁBAL URIBE, pertenecientes al Departamento de Ciencias Químicas y Recursos Naturales de la Universidad de La Frontera, y es presentada para su revisión por los miembros de la comisión examinadora. FELIPE EDUARDO MORAGA NICOLÁS Dr. Andrés Quiroz Director programa de Doctorado en Dra. Ana Mutis T. Ciencias de Recursos Naturales Dr. Emilio Hormazábal U. Dr. Patricio Iturriaga V. Dra. Mónica Rubilar Directora Académica de Postgrado Universidad de La Frontera Dr. Andrés Quiroz C. Dr. Alejandro Urzúa M. Dr. Leonardo Guzmán G. Dedico esta tesis a mi Madre por su apoyo amor y confianza Agradecimientos/Acknowledgments Este escrito no solo refleja los resultados de una investigación, sino también, interés y aprecio por los productos naturales. El desarrollo de este trabajo no habría sido posible sin el apoyo de todas aquellas personas, que directa o indirectamente, me han permitido llevar a cabo los objetivos propuestos en esta investigación. De este modo, las siguientes líneas expresan mis agradecimientos más sinceros a quienes colaboraron en hacer esto una realidad. -

Town of Jupiter
TOWN OF JUPITER DATE: November 19, 2019 TO: Honorable Mayor and Members of Town Council THRU: Matt Benoit, Town Manager FROM: David Brown, Utilities Director MB John R. Sickler, Director of Planning and Zoning SUBJECT: Glyphosate Use Reduction –Resolution to call for a reduction in the use of products containing glyphosate by the Town and its contractors and encouraging a reduction in use by the public HEARING DATES: ETF 11/4/19 PZ #19-4030 TC 11/19/19 Resolution #108-19 EXECUTIVE SUMMARY: Consideration of a resolution recognizing the potential human health and environmental benefits of reducing the use of glyphosate-based herbicides by Town employees and its contractors. Background While glyphosate and formulations such as Roundup have been approved by regulatory bodies worldwide, concerns about their effects on humans and the environment persist, and have grown as the global usage of glyphosate increases. There is a growing belief by some that glyphosate may be carcinogenic. Much of this concern is related to use on food crops and direct exposure via application of the herbicide. In 2015, glyphosate was classified as a probable carcinogen by the International Agency for Research on Cancer, an arm of the World Health Organization (Attachment A). However, this designation was not without controversy (Attachment B) and it is important to note that the U.S. Environmental Protection Agency (EPA) maintains that glyphosate is not likely to be carcinogenic to humans and is not currently banned for use by the U.S. government (pg. 143, Attachment C). In addition, the University of Florida Institute of Food and Agricultural Sciences continues to recommend the use of glyphosate as a weed control tool with the caveat that users of these products must carefully read and follow all label directions (Attachment D). -
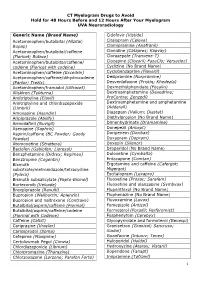
CT Myelogram Drugs to Avoid Hold for 48 Hours Before and 12 Hours After Your Myelogram UVA Neuroradiology
CT Myelogram Drugs to Avoid Hold for 48 Hours Before and 12 Hours After Your Myelogram UVA Neuroradiology Generic Name (Brand Name) Cidofovir (Vistide) Acetaminophen/butalbital (Allzital; Citalopram (Celexa) Bupap) Clomipramine (Anafranil) Acetaminophen/butalbital/caffeine Clonidine (Catapres; Kapvay) (Fioricet; Butace) Clorazepate (Tranxene-T) Acetaminophen/butalbital/caffeine/ Clozapine (Clozaril; FazaClo; Versacloz) codeine (Fioricet with codeine) Cyclizine (No Brand Name) Acetaminophen/caffeine (Excedrin) Cyclobenzaprine (Flexeril) Acetaminophen/caffeine/dihydrocodeine Desipramine (Norpramine) (Panlor; Trezix) Desvenlafaxine (Pristiq; Khedezla) Acetaminophen/tramadol (Ultracet) Dexmethylphenidate (Focalin) Aliskiren (Tekturna) Dextroamphetamine (Dexedrine; Amitriptyline (Elavil) ProCentra; Zenzedi) Amitriptyline and chlordiazepoxide Dextroamphetamine and amphetamine (Limbril) (Adderall) Amoxapine (Asendin) Diazepam (Valium; Diastat) Aripiprazole (Abilify) Diethylpropion (No Brand Name) Armodafinil (Nuvigil) Dimenhydrinate (Dramamine) Asenapine (Saphris) Donepezil (Aricept) Aspirin/caffeine (BC Powder; Goody Doripenem (Doribax) Powder) Doxapram (Dopram) Atomoxetine (Strattera) Doxepin (Silenor) Baclofen (Gablofen; Lioresal) Droperidol (No Brand Name) Benzphetamine (Didrex; Regimex) Duloxetine (Cymbalta) Benztropine (Cogentin) Entacapone (Comtan) Bismuth Ergotamine and caffeine (Cafergot; subcitrate/metronidazole/tetracycline Migergot) (Pylera) Escitalopram (Lexapro) Bismuth subsalicylate (Pepto-Bismol) Fluoxetine (Prozac; Sarafem)