FORESTS and GENETICALLY MODIFIED TREES FORESTS and GENETICALLY MODIFIED TREES
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Backgrounder: the New York Botanical Garden's Legacy Of
Moore in America: Monumental Sculpture at The New York Botanical Garden May 24 – November 2, 2008 Backgrounder: The New York Botanical Garden’s Legacy of Natural and Designed Landscapes The New York Botanical Garden, a 250-acre site that has been designated a National Historic Landmark, offers a wealth of beautiful landscapes, including a hardwood Forest, ponds, lakes, streams, rolling hills with dramatic rock outcroppings carved by glaciers, and New York City’s only freshwater river, which runs through the heart of the Forest in a magnificent rock gorge. These picturesque natural features have been further enhanced by more than a century of artful plantings, gardens, and landscapes designed by the nation’s leading landscape architects and garden designers. As a result of both its natural and human legacies, the Botanical Garden today offers an exceptional setting for outdoor sculpture. Scenic beauty and stunning natural features Following the New York State Governor’s approval on April 28, 1891, of The New York Botanical Garden Act of Incorporation, a site needed to be selected for the location of this new educational and scientific institution. Selection turned to an undeveloped park in the central Bronx. In 1887, a published description of this area notes, “it would be difficult to do justice to the exquisite loveliness of this tract without seeming to exaggerate…gigantic trees, centuries old, crown these summits, while great moss and ivy-covered rocks project here and there at different heights above the surface of the water, increasing the wildness of the science.” An 1893 newspaper account describes the romantic vistas of an old stone house, snuff mill, and other artifacts of previous land use, while surrounded with “almost every tree known to the American forest in the Northern clime.” The underlying bedrock, dark gray Fordham gneiss, shapes many rock outcrops, rolling hills, and steep slopes, ranging from 20 to 180 feet above sea level. -

Urban Agriculture
GSDR 2015 Brief Urban Agriculture By Ibrahim Game and Richaela Primus, State University of New York College of Forestry and Environmental Science Related Sustainable Development Goals Goal 01 End poverty in all its forms everywhere (1.1, 1.4, 1.5 ) Goal 02 End hunger, achieve food security and improved nutrition and promote sustainable agriculture (2.1, 2.3, 2.4, 2.c) Goal 12 Ensure sustainable consumption and production patterns (12.1, 12.2, 12.3, 12.4,12.5, 12.7, 12.8) Goal 15 Protect, restore and promote sustainable use of terrestrial ecosystems, sustainably manage forests, combat desertification, and halt and reverse land degradation and halt biodiversity loss (15.9 ) *The views and opinions expressed are the authors’ and do not represent those of the Secretariat of the United Nations. Online publication or dissemination does not imply endorsement by the United Nations. Authors can be reached at [email protected] and [email protected]. Introduction Examples of UEA include community gardens, vegetable gardens and rooftop farms, which exist Urban Agriculture (UA) and peri-urban agriculture can worldwide and are playing important roles in the urban be defined as the growing, processing, and distribution food systems. 17 CEA includes any form of agriculture of food and other products through plant cultivation where environmental conditions (such as, light, and seldom raising livestock in and around cities for temperature, humidity, radiation and nutrient cycling) 1 2 feeding local populations. Over the last few years, are controlled in conjunction with urban architecture UA has increased in popularity due to concerns about or green infrastructure. -

Biohazardous Waste Treatment
Biological Safety: Principles & Applications for Lab Personnel Presented by: Biological Safety Office http://biosafety.utk.edu Introduction OVERVIEW OF BIOSAFETY PROGRAM Introduction to Laboratory Safety When entering the laboratory environment you are likely to contact hazards that you would not encounter on a daily basis. These can include: • Physical hazards • Chemical hazards • Radiological hazards • Biological hazards Biohazards Any biological agent or condition that poses a threat to human, animal, or plant health, or to the environment. Examples include, but not limited to: • Agents causing disease in humans, animals or plants (bacteria, viruses, etc.) • Toxins of biological origin • Materials potentially containing infectious agents or biohazards .Blood, tissues, body fluids etc. .Waste, carcasses etc. • Recombinant DNA (depends) • Nanoparticles w/biological effector conjugates (depends) Types of Biohazards: What they are.... How they’ll look.... Regulations & Standards The Institutional Biosafety Committee Institutional Biosafety Committees (IBC) are required by the National Institutes of Health Office of Biological Activities (NIH; OBA) for institutions that receive NIH funding and conduct research using recombinant or synthetic nucleic acids. The UT IBC is composed of 14 UT-affiliated members and 2 non-affiliated members, who collectively offer a broad range of experience in research, safety and public health. Functions of the IBC Projects requiring IBC review • Performing both initial and annual • Those involving recombinant -

Agroforestry : Fact-Sheet
Agroforestry fact sheet Agroforestry is the mixture of trees and crops in cultivated parcels. It was a traditional system in both tropical and temperate countries before agriculture intensification during the last century. Very recent results show that it may be a key option for the future of modern agriculture, including in temperate countries. 10 years ago, agroforestry was still ignored by farmers, foresters, policy makers of developed coun- tries. But times have changed. Agroforestry is an utmost example of ecological intensification : it in- creases land productivity and offers at the same time many environmental services. A French national scheme for planting half a million hectares of agroforestry during the next 25 years was based on res- ults obtained by INRA, Montpellier. A walnut – wheat agroforestry plot at the Restinclières farm, tree pruning time (left) Monitoring water sap flow through tap roots and superficial roots to document the hydraulic lift on walnut agroforestry trees (right) Increase of productivity with agroforestry The mix of trees and crops is the fundamental principle of agroforestry. INRA researchers showed that the production from one hectare of a walnut/wheat mix is the same as for 1.4 hectares with trees and crops separated. This is a 40% increase in productivity, far better than any other innovation intro- duced by agronomists in the recent past. This is achieved with an optimal 50 to 100 trees/hectare dens- ity, and an appropriate management of the system (tree line orientation, tree pruning regime, winter crops). Agroforestry trees grow very fast, faster than trees in forest plantations. Rightly associated and appropriately managed, trees and annual crops establish a synergy in the use of the vital resources of light, water and soil nutrients. -

Overview of the Dallas Arboretum and Botanical Garden the Mission
Overview of The Dallas Arboretum and Botanical Garden The Mission Our mission makes us much more than just a beautiful place as we are charged to: Provide a place for the art and enjoyment of horticulture Provide for the education of adults and children Provide research to return to the field Do so in a fiscally responsible way 2 DALLAS ARBORETUM The Jonsson Color Garden 3 DALLAS ARBORETUM The Palmer Fern Dell 4 DALLAS ARBORETUM The Paseo de Flores 5 DALLAS ARBORETUM A Woman’s Garden Phase One 6 DALLAS ARBORETUM A Woman’s Garden Phase Two 7 DALLAS ARBORETUM The McCasland Sunken Garden 8 DALLAS ARBORETUM The Boswell Family Garden 9 DALLAS ARBORETUM Nancy’s Garden 10 DALLAS ARBORETUM The Rose Mary Haggar Rose Garden 11 DALLAS ARBORETUM The Nancy Clements Seay Magnolia Glade 12 DALLAS ARBORETUM The Martha Brooks Camellia Garden 13 DALLAS ARBORETUM The Nancy Rutchik Red Maple Rill 14 DALLAS ARBORETUM The Martin Rutchik Concert Stage and Lawn 15 DALLAS ARBORETUM The Lay Family Garden 16 DALLAS ARBORETUM The Henry Lindsley Shadow Garden 17 DALLAS ARBORETUM The Water Wise Garden 18 DALLAS ARBORETUM Artscape, Fine Art Show and Sale 19 DALLAS ARBORETUM A Tasteful Place Opened Fall 2017 A Two and a Half Acre Fruit, Herb, and Vegetable Garden Teaching Visitors How to Grow Local and Sustainable Produce and Cook in Nutritious Ways. Area for tastings or demonstrations each day. An enclosed building for cooking classes and lectures. Four quadrants with plantings in trays that are moved to the greenhouse when dormant. Orchard and vineyard areas. -

Field Trials for GM Food Get Green Light in India
NATURE|Vol 448|23 August 2007 NEWS IN BRIEF Field trials for GM food Asthmatics win payment in diesel-fumes lawsuit get green light in India Asthma patients in Tokyo last week welcomed a cash settlement India’s first genetically modified (GM) food from car manufacturers and crop is a step closer to reaching the dining the Japanese government. The table. The government has approved field one-time payment resolved a trials for a strain of brinjal (aubergine) decade-long legal battle in which carrying a Bt (Bacillus thuringiensis) gene. the asthmatics blamed diesel car Jalna-based Mahyco, the Indian venture fumes for their illness. KIM KYUNG-HOON/REUTERS of US seed giant Monsanto, claims its insect- The automakers, including resistant variety gives better yields with Toyota, Honda and Nissan, will less pesticide use. To avoid possible cross- provide ¥1.2 billion (US$10.5 contamination with farmers’ crops, the trials million) to the plaintiffs, and a will be carried out in government farms. further ¥3.3 billion to support But critics already campaigning against Bt a five-year health plan for the cotton — currently the only GM crop grown patients. The central government in India — say the brinjal trials are illegal. and Tokyo metropolitan Full biosafety data on the brinjal tests government will each contribute ¥6 billion to the medical programme. have not yet been generated, says Kavitha Kuruganti of the Centre for Sustainable scientists — to pay for relocation, housing starting dose “should be considered for Agriculture, based in Hyderabad: “The trials and research. “This is about the rescue of patients with certain genetic variations”. -

Synthesizing Ecosystem Implications of Mistletoe Infection
Environmental Research Letters LETTER • OPEN ACCESS Related content - Networks on Networks: Water transport in Mistletoe, friend and foe: synthesizing ecosystem plants A G Hunt and S Manzoni implications of mistletoe infection - Networks on Networks: Edaphic constraints: the role of the soil in vegetation growth To cite this article: Anne Griebel et al 2017 Environ. Res. Lett. 12 115012 A G Hunt and S Manzoni - Impact of mountain pine beetle induced mortality on forest carbon and water fluxes David E Reed, Brent E Ewers and Elise Pendall View the article online for updates and enhancements. This content was downloaded from IP address 137.154.212.215 on 17/12/2017 at 21:57 Environ. Res. Lett. 12 (2017) 115012 https://doi.org/10.1088/1748-9326/aa8fff LETTER Mistletoe, friend and foe: synthesizing ecosystem OPEN ACCESS implications of mistletoe infection RECEIVED 28 June 2017 Anne Griebel1,3 ,DavidWatson2 and Elise Pendall1 REVISED 1 Hawkesbury Institute for the Environment, Western Sydney University, Locked Bag 1797, Penrith, NSW, Australia 12 September 2017 2 Institute for Land, Water and Society, Charles Sturt University, PO box 789, Albury, NSW, Australia ACCEPTED FOR PUBLICATION 3 Author to whom any correspondence should be addressed. 29 September 2017 PUBLISHED E-mail: [email protected] 16 November 2017 Keywords: mistletoe, climate change, biodiversity, parasitic plants, tree mortality, forest disturbance Original content from this work may be used Abstract under the terms of the Creative Commons Biotic disturbances are affecting a wide range of tree species in all climates, and their occurrence is Attribution 3.0 licence. contributing to increasing rates of tree mortality globally. -
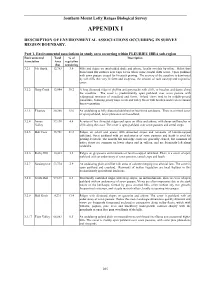
A Biological Survey of the Southern Mount Lofty Ranges
Southern Mount Lofty Ranges Biological Survey APPENDIX I DESCRIPTION OF ENVIRONMENTAL ASSOCIATIONS OCCURRING IN SURVEY REGION BOUNDARY. Part 1. Environmental associations in study area occurring within FLEURIEU IBRA sub-region Environmental Total % of Description Association Area vegetation (ha) remaining 3.2.1 Mt. Rapid 12,763 3.9 Hills and ridges on interbedded shale and arkose, locally overlain by tillite. Relict fans form broad flat surfaces near Cape Jervis where some coastal cliffs occur. Open parkland with sown pasture is used for livestock grazing. The scenery of the coastline is dominated by tall cliffs that vary in form and steepness, the amount of rock outcrop and vegetative cover. 3.2.2 Deep Creek 12,984 30.2 A long dissected ridge of phyllite and greywacke with cliffs, or beaches and dunes along the coastline. The cover is predominantly open parkland over sown pasture with widespread remnants of woodland and forest. Inland views tend to be middle-ground panoramic, featuring grassy ridge crests and valley floors with bracken and reed or remnant forest vegetation. 3.2.3 Fleurieu 30,389 15.6 An undulating to hilly dissected tableland on lateritized sandstone. There is a mixed cover of open parkland, forest plantation and woodland. 3.2.4 Inman 37,130 4.4 A series of low dissected ridges and spurs on tillite and arkose, with dunes and beaches or Valley cliffs along the coast. The cover is open parkland over sown pastures and cereal crops. 3.2.5 Bob Tiers 15,761 21.3 Ridges on schist and gneiss with dissected slopes and remnantsof laterite-capped tableland. -

Agriculture and the Future of Food: the Role of Botanic Gardens Introduction by Ari Novy, Executive Director, U.S
Agriculture and the Future of Food: The Role of Botanic Gardens Introduction by Ari Novy, Executive Director, U.S. Botanic Garden Ellen Bergfeld, CEO, American Society of Agronomy, Crop Science Society of America, and Soil Science Society of America The more than 320 million Americans alive today depend on plants for our food, clothing, shelter, medicine, and other critical resources. Plants are vital in today’s world just as they were in the lives of the founders of this great nation. Modern agriculture is the cornerstone of human survival and has played extremely important roles in economics, power dynamics, land use, and cultures worldwide. Interpreting the story of agriculture and showcasing its techniques and the crops upon which human life is sustained are critical aspects of teaching people about the usefulness of plants to the wellbeing of humankind. Botanical gardens are ideally situated to bring the fascinating story of American agriculture to the public — a critical need given the lack of exposure to agricultural environments for most Americans today and the great challenges that lie ahead in successfully feeding our growing populations. Based on a meeting of the nation’s leading agricultural and botanical educators organized by the U.S. Botanic Garden, American Society of Agronomy, Crop Science Society of America, and Soil Science Society of America, this document lays out a series of educational narratives that could be utilized by the U.S. Botanic Garden, and other institutions, to connect plants and people through presentation -

Jacksonville Arboretum & Botanical Gardens Receives Grants to Start
Jacksonville Arboretum & Botanical Gardens e-Newsletter February 2021 EDITION Hours: 8 a.m. to 5 p.m. Jacksonville Arboretum & Botanical Gardens Receives Grants to Start Master Plan Process Jacksonville, Fla. (Feb. 10, 2021) – Later this month, the Arboretum will begin the exciting process of developing a master plan to design and install botanical gardens on the property. The master plan process was made possible by generous grants of $30,000 from the Delores Barr Weaver Legacy Fund and $10,000 from the River Branch Foundation. The planning process will take about eight months to complete. The addition of botanical gardens is the latest in a continuous plan to propel the Arboretum into a best-in-class destination in the southeast. Executive Director Dana Doody noted that each project is being planned carefully to add as much value as possible while managing the non-profit’s budget plan in unprecedented times. In addition to the grant for the master plan design, the Delores Barr Weaver Legacy Fund also awarded the Arboretum a $70,000 challenge grant for the first phase of the implementation. The 1:1 challenge grant will launch a community campaign following the completion of the design plan. “The Arboretum is an important community asset which brings thousands of people to its special trails, many of which are ADA accessible,” said Delores Barr Weaver. “The plan will provide a vision for a botanical destination, sure to benefit our citizens for years to come.” The process will take into account the Arboretum’s unique qualities, Florida’s seasons and Jacksonville’s ecosystems and native horticulture. -

Course Handout for Introduction to Forest Gardening
COURSE HANDOUT FOR INTRODUCTION TO FOREST GARDENING Complied by Jess Clynewood and Rich Wright Held at Coed Hills Rural Art Space 2010 ETHICS AND PRINCIPLES OF PERMACULTURE Care for the Earth v Care for the people v Fair shares PRINCIPLES Make the least change for the greatest effect v Mistakes are tools for learning v The only limits to the yield of a system are imagination and understanding Observation – Protracted and thoughtful observation rather than prolonged and thoughtless action. Observation is a key tool to re-learn. We need to know what is going on already so that we don’t make changes we will later regret. Use and value diversity - Diversity allows us to build a strong web of beneficial connections. Monocultures are incredibly fragile and prone to pests and diseases – diverse systems are far more robust and are intrinsically more resilient. Relative Location and Beneficial Connections – View design components not in isolation but as part of a holistic system. Place elements to maximise their potential to create beneficial connections with other elements. Multi-functional Design – Try and gain as many yields or outputs from each element in your design as possible. Meet every need in multiple ways, as many elements supporting each important function creates stability and resilience. Perennial systems – minimum effort for maximum gain Create no waste - The concept of waste is essentially a reflection of poor design. Every output from one system could become the input to another system. We need to think cyclically rather than in linear systems. Unmet needs = work, unused output = pollution. Stacking – Make use of vertical as well as horizontal space, filling as many niches as possible. -

Influence of Coarse Woody Debris on Seedlings and Saplings in A
INFLUENCE OF COARSE WOODY DEBRIS ON SEEDLINGS AND SAPLINGS IN A PINUS PALUSTRIS WOODLAND by ALEXANDRA LOGAN JUSTIN L. HART, COMMITTEE CHAIR MATTHEW C. LAFEVOR ARVIND A.R. BHUTA A THESIS Submitted in partial fulfillment of the requirements for the degree of Master of Science in the Department of Geography in the Graduate School of The University of Alabama TUSCALOOSA, ALABAMA 2020 Copyright Alexandra Logan 2020 ALL RIGHTS RESERVED 2 ABSTRACT Coarse woody debris (CWD) has beneficial effects on plant growth and establishment. Longleaf pine (Pinus palustris Mill.) stands support relatively low amounts of CWD — 2 to 30 m3 ha-1. In April 2011, an EF3 tornado passed through the Oakmulgee Ranger District of the Talladega National Forest in the Fall Line Hills of Alabama. This disturbance resulted in the large addition of CWD to a longleaf pine woodland, and a rare opportunity to analyze how CWD can influence a managed, pine woodland. The goal of this study was to examine the effect of CWD on woody plant richness, density, and growth rate (quantified by height) in a longleaf pine woodland that experienced a catastrophic wind disturbance. A total of three 1 m2 quadrats were established against either side of a piece of CWD (> 3 m in length and ≥ 10 cm in diameter). Another quadrat was established at least 3 m away from the focal CWD piece. For each plot, the presence and height of every woody plant (< 5 cm dbh) were recorded. Sapling density, oak and hickory density, and organic matter were all found to be significantly higher in quadrats adjacent to CWD than away (all p < 0.05).