Those on the Latter by Katayama (I) and Minamibuchi (I)
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Geological Record of Prehistoric Tsunamis in Mugi Town, Facing The
Shimada et al. Progress in Earth and Planetary Science (2019) 6:33 Progress in Earth and https://doi.org/10.1186/s40645-019-0279-9 Planetary Science RESEARCH ARTICLE Open Access Geological record of prehistoric tsunamis in Mugi town, facing the Nankai Trough, western Japan Yumi Shimada1,3* , Shigehiro Fujino2, Yuki Sawai3, Koichiro Tanigawa3, Dan Matsumoto3, Arata Momohara4, Megumi Saito-Kato5, Masaki Yamada6, Eri Hirayama1, Takahiro Suzuki1 and Catherine Chagué7 Abstract Stratigraphic and paleontological investigations in Mugi Town, on the Pacific coast of Shikoku Island, revealed evidence of as many as five tsunami inundations from events along the Nankai Trough between 5581 and 3640 cal yr BP. Nine event deposits (E1–E9) were identified in cores ranging in length from 2 to 6 m, consisting of sandy and gravelly layers interbedded with organic-rich mud. Sedimentary structures in the event deposits observed by computed tomography included normal grading and sharp lower stratigraphic contacts. Event deposits E3, E6, E7, and E8 contained mainly brackish-marine diatom species, suggesting that they had been deposited during inundation by seawater. In addition, fossil diatom assemblages were markedly different above and below event deposits E3, E4, E6, and E8. For example, assemblages below event deposit E6 were dominated by a freshwater species (Ulnaria acus), whereas assemblages above it were predominantly brackish-marine (Diploneis smithii, Fallacia forcipata,andFallacia tenera). We attributed these changes to the increase of marine influence due to coastal subsidence associated with subduction-zone earthquakes, as documented in the 1946 Showa-Nankai earthquakes. We conclude that event deposits E3, E6, and E8 and perhaps E4 and E7 were deposited by tsunamis generated by subduction zone earthquakes along the Nankai Trough. -

Genetic Lineage of the Amami Islanders Inferred from Classical Genetic Markers
bioRxiv preprint doi: https://doi.org/10.1101/2021.04.18.440379; this version posted April 19, 2021. The copyright holder for this preprint (which was not certified by peer review) is the author/funder, who has granted bioRxiv a license to display the preprint in perpetuity. It is made available under aCC-BY-NC-ND 4.0 International license. Genetic lineage of the Amami islanders inferred from classical genetic markers Yuri Nishikawa and Takafumi Ishida Department of Biological Sciences, Graduate School of Science, The University of Tokyo, Tokyo, Japan Correspondence: Yuri Nishikawa, Department of Biological Sciences, Graduate School of Science, The University of Tokyo, Hongo 7-3-1, Bunkyo-ku, Tokyo 113-0033, Japan. E-mail address: [email protected] 1 bioRxiv preprint doi: https://doi.org/10.1101/2021.04.18.440379; this version posted April 19, 2021. The copyright holder for this preprint (which was not certified by peer review) is the author/funder, who has granted bioRxiv a license to display the preprint in perpetuity. It is made available under aCC-BY-NC-ND 4.0 International license. Abstract The peopling of mainland Japan and Okinawa has been gradually unveiled in the recent years, but previous anthropological studies dealing people in the Amami islands, located between mainland Japan and Okinawa, were less informative because of the lack of genetic data. In this study, we collected DNAs from 104 subjects in two of the Amami islands, Amami-Oshima island and Kikai island, and analyzed the D-loop region of mtDNA, four Y-STRs and four autosomal nonsynonymous SNPs to clarify the genetic structure of the Amami islanders comparing with peoples in Okinawa, mainland Japan and other regions in East Asia. -

Local Dishes Loved by the Nation
Sapporo 1 Hakodate 2 Japan 5 3 Niigata 6 4 Kanazawa 15 7 Sendai Kyoto 17 16 Kobe 10 9 18 20 31 11 8 ocal dishes Hiroshima 32 21 33 28 26 19 13 Fukuoka 34 25 12 35 23 22 14 40 37 27 24 29 Tokyo loved by 41 38 36 Nagoya 42 44 39 30 Shizuoka Yokohama 43 45 Osaka Nagasaki 46 Kochi the nation Kumamoto ■ Hokkaido ■ Tohoku Kagoshima L ■ Kanto ■ Chubu ■ Kansai 47 ■ Chugoku ■ Shikoku Naha ■ Kyushu ■ Okinawa 1 Hokkaido 17 Ishikawa Prefecture 33 Okayama Prefecture 2 Aomori Prefecture 18 Fukui Prefecture 34 Hiroshima Prefecture 3 Iwate Prefecture 19 Yamanashi Prefecture 35 Yamaguchi Prefecture 4 Miyagi Prefecture 20 Nagano Prefecture 36 Tokushima Prefecture 5 Akita Prefecture 21 Gifu Prefecture 37 Kagawa Prefecture 6 Yamagata Prefecture 22 Shizuoka Prefecture 38 Ehime Prefecture 7 Fukushima Prefecture 23 Aichi Prefecture 39 Kochi Prefecture 8 Ibaraki Prefecture 24 Mie Prefecture 40 Fukuoka Prefecture 9 Tochigi Prefecture 25 Shiga Prefecture 41 Saga Prefecture 10 Gunma Prefecture 26 Kyoto Prefecture 42 Nagasaki Prefecture 11 Saitama Prefecture 27 Osaka Prefecture 43 Kumamoto Prefecture 12 Chiba Prefecture 28 Hyogo Prefecture 44 Oita Prefecture 13 Tokyo 29 Nara Prefecture 45 Miyazaki Prefecture 14 Kanagawa Prefecture 30 Wakayama Prefecture 46 Kagoshima Prefecture 15 Niigata Prefecture 31 Tottori Prefecture 47 Okinawa Prefecture 16 Toyama Prefecture 32 Shimane Prefecture Local dishes loved by the nation Hokkaido Map No.1 Northern delights Iwate Map No.3 Cool noodles Hokkaido Rice bowl with Tohoku Uni-ikura-don sea urchin and Morioka Reimen Chilled noodles -

Please Read This Book Before Coming to Japan
表4 表1 JET プログラム参加者用ハンドブック Please read this book before coming to Japan. 来日前に必ずこの冊子を読んでください。 撮影地 新倉山浅間公園(山梨県富士吉田市) Arakurayama Sengen Park (Yamanashi Prefecture Fujiyoshida City) Designed by Amari JET プログラム参加者情報・JET Programme Participant Information 名前 Name JET 番号 JET No. 勤務先情報・Workplace Information 任用団体 Contracting Organisation 住所 Address 電話番号 FAX 番号 Telephone No. FAX No. E メール E-mail 担当者氏名 Supervisor’s Name 緊急通報用電話番号・Emergency Phone Numbers 警察 Police ・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・ 110 救急車/消防 Ambulance/Fire ・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・ 119 警察相談ダイヤル Police Consultation Centre ・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・ #9110 緊急通報時に役立つフレーズ・Emergency Phrases JET プログラムの Web ページにはたくさんの情報があります 警察・Police Tasukete kudasai! (Please help!) Useful information can be found on the JET Programme’s website. ________________ ga nusumaremashita. (My_________________has been stolen.) Jūsho wa _________________ desu. (My address is _________________.) Namae wa _________________desu. (My name is _________________.) CLAIR ニュースはかならずよみましょう Denwa bangō wa___________ desu. (My telephone number is ____________.) 救急車/消防・Ambulance/Fire Please be sure to always read CLAIR News. Kaji desu! (Fire!) Jiko desu. (There has been an accident.) Kyūkyūsha wo onegai shimasu. (Send an ambulance, please.) Jūsho wa _________________ desu. (My address is _________________.) ** CLAIR News is a newsletter in Japanese and English that is e-mailed to current JET Namae wa _________________desu. (My name is _________________.) participants -

Wood Use Promotion Division Forestry Agency Preface
Wood Use Promotion Division Forestry Agency Preface As forest resources in Japan have come to maturity, it is important to promote cyclical use of them through increasing the use of lumber. Besides the housing sector, the use of lumber is expected to grow in public facilities, office buildings and others. Outside Japan, demand for lumber is forecast to increase due to economic development and population growth in emerging economies. Japanese lumber is expected to meet such demand as well. The Ministry of Agriculture, Forestry and Fisheries is promoting programs to reinforce exports of agricultural, forest and fishery products with the 1-trillion-yen target set for them in the Export Promotion Strategy of Agriculture, Forestry, Fisheries and Food Products it adopted in fiscal 2013. Exports of forest products (lumber and special forest products) increased to 27.4 billion yen in fiscal 2016. But there remain challenges with them as logs account for a large portion of exports , and the number of export destinations is limited. In the meantime, efforts to boost exports of forest products are emerging in various parts of Japan. Among them is the production of highly durable lumber treated at high temperatures and high-performance wooden window products. The Forestry Agency, in cooperation with prefectural governments, has collected such efforts underway to increase exports of forest products in various parts of the country and compiled this report as the “Case Studies of Forest Products Exports – Japanese Lumber to the World”. We hope this report will serve as a reference for efforts to increase exports of lumber and forest products. -

Regional Social System for Specialized Medical Care in Hematologic Malignancies: a Pilot Study
ORIGINAL RESEARCH Regional social system for specialized medical care in hematologic malignancies: a pilot study M Takita 1, Y Tanaka 1, T Matsumura 1, Y Kishi 1, Y Kodama 1, T Nishimura 1, T Goto 2, M Nagai 3, M Kami 1 1Division of Social Communication System for Advanced Clinical Research, The Institute of Medical Science, The University of Tokyo, Tokyo, Japan 2Division of Hematology, Tokushima Red Cross Hospital, Komatsushima-city, Tokushima, Japan 3Tokushima Prefectural Central Hospital, Tokushima-city, Tokushima, Japan Submitted: 24 October 2008; Resubmitted: 3 March 2009; Published: 28 July 2009 Takita M, Tanaka Y, Matsumura T, Kishi Y, Kodama Y, Nishimura T, Goto T, Nagai M, Kami M Regional social system for specialized medical care in hematologic malignancies: a pilot study Rural and Remote Health 9: 1106. (Online), 2009 Available from: http://www.rrh.org.au A B S T R A C T Introduction: The uneven distribution of physicians in Japan disadvantages rural and remote patients with hematological malignancies to the extent that they may not receive standard treatments. There are 7 core regional medical centers in Tokushima Prefecture. A Tokushima rural medical center’s clinical hematology division experienced difficulty in treating patients due to a lack of physicians despite an increasing number of patients with hematological malignancies. The aim of this pilot study was to investigate the regional medical supply and demand associated with newly diagnosed hematological malignancies in Tokushima Prefecture, Japan. Methods: The study investigated the home addresses of patients with newly diagnosed acute leukemia, malignant lymphoma and multiple myeloma who were hospitalized in the 7 core Tokushima centers in 2006. -
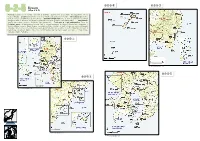
6-2-5-④ 6-2-5-③ Kyusyu 6-2-5 (Map 6-2-5) Province: Kumamoto Pref
6-2-5-④ 6-2-5-③ Kyusyu 6-2-5 (Map 6-2-5) Province: Kumamoto Pref. at west, Oita Pref. at northeast, Miyazaki Pref. at southeast, and Kagoshima Pref. at south of Kyushu Location: Kyushu lies at west of Shikoku and southwest of Honshu Air temperature: 17.8˚C (annual average, at Ushibuka City, Kumamoto.) Seawater temperature: 22.9˚C, 22.2 ˚C and 20.9 ˚C (annual average, at east off Aburatsu, southwest off Kushikijima (Is.) and Yatsushiro Sea, respectively) Precipitation: 2,027.9 mm (annual average, Ushibuka City, Kumamoto) Total area of coral communities: 581.8 ha Protected areas: Unzen-Amakusa National Park: at around Amakusa, including 3 Marine Park Zones and 2 Protected Water Surfaces; Nippo Kaigan Quasi-National Park: coastline at south of Oita and north of Miyazaki, including 2 Marine Park Zones; Nichinan Kaigan Quasi-National Park: coastline at south of Miyazaki and east of Kagoshima, including 1 Marine Park Zone; Kirishima-Yaku National Park: a part of coastline in Kagoshima, including 2 Marine Park Zones. 6-2-5-① *“号”on this map means“site”. 6-2-5-⑤ 6-2-5-② *“号”on this map means“site”. 6-2-5-② 6-2-5-① *“号”on this map means“site”. 6-2-5-④ 6-2-5-③ *“号”on this map means“site”. 6-2-5-⑤ 06 Coral Reefs of Japan a. Kumamoto Prefecture (Map 6-2-5-①) Satoshi Nojima 1 Corals and coral reefs Photo. 1. Tabulate Acropora dominant community in Kuwashima 1. Geographical features (Is.), Ushibuka City, Kumamoto Prefecture. On the west of Kyushu in Kumamoto Prefecture lie the Amakusa Islands. -

VISIT JAPAN Ambassadors
“Visit Japan” Ambassadors Fifty-five Ambassadors as of February 22, 2017 (listed in the order of the Japanese syllabary) Takanori ADACHI [Director, Adachi Museum of Art] Maintaining Japan’s finest gardens to promote the dissemination of Japanese culture; creating a welcoming environment for international visitors The Adachi Museum of Art in Yasugi, Shimane Prefecture, has large Japanese gardens. These perfectly designed gardens are meticulously maintained, and their beauty awes international visitors. For the 11 years from 2003*, the museum was chosen as having the finest Japanese gardens by the U.S. magazine The Journal of Japanese Gardening. The gardens at the museum have been promoting the dissemination of Japanese culture as well as drawing increasing numbers of international visitors. The museum began efforts to attract more visitors from overseas earlier than other museums. In 1992, it began offering discounted admissions fees to international visitors. In 2006, it was certified by the “Visit Japan” Information Network as Japan’s first art museum to double as a tourist information center. *as of January 2014 Daishiro ARAMAKI [President, Kabukizabutai] Promoting the appeal of Japanese culture overseas through Kabuki; improving the environment for international audiences Kabuki is a major Japanese performing art. While serving as director of Shochiku Co., Ltd., Mr. Aramaki led a corporate effort to internationalize Kabuki by supporting Kabuki performances overseas and helping to hold Kabuki performances in English. In this endeavour, Kabuki actors have visited 108 municipalities of 35 countries, giving Kabuki performances more than 60 times to date. Each Kabuki performance attracted a large audience of several thousand to tens of thousands of people, helping to increase the number of Kabuki enthusiasts around the world. -

A Bird's Eye View of Okinawa
A Bird’s Eye View of Okinawa by HIH Princess Takamado, Honorary President ne of the most beautiful of the many O“must visit” places in Japan is the Ryukyu Archipelago. These islands are an absolute treasure trove of cultural, scenic and environmental discoveries, and the local people are known for their warmth and welcoming nature. Ikebana International is delighted to be able to host the 2017 World Convention in Okinawa, and I look forward to welcoming those of you who will be joining us then. 13 Kagoshima Kagoshima pref. Those who are interested in flowers are generally interested in the environment. In many cultures, flowers and birds go together, and so, Osumi Islands Tanega too, in my case. As well as being the Honorary President of Ikebana International, I am also the Yaku Honorary President of BirdLife International, a worldwide conservation partnership based in Cambridge, UK, and representing approximately 120 countries or territories. In this article, I Tokara Islands would like introduce to you some of the birds of Okinawa Island as well as the other islands in the Ryukyu Archipelago and, in so doing, to give you Amami a sense of the rich ecosystem of the area. Amami Islands Kikaiga One Archipelago, Six Island Tokuno Groups The Ryukyu Archipelago is a chain of islands Okinawa pref. Okino Erabu that stretches southwest in an arc from Kyushu (Nansei-shoto) to Chinese Taiwan. Also called the Nansei Islands, the archipelago consists of over 100 islands. Administratively, the island groups of Kume Okinawa Naha Osumi, Tokara and Amami are part of Kagoshima Prefecture, whilst the island groups Ryukyu Archipelago of Okinawa, Sakishima (consisting of Miyako Okinawa Islands and Yaeyama Islands), Yonaguni and Daito are part of Okinawa Prefecture. -

Ratio of Elderly Population Who Are Living Alone, Disabled, and Cared in Nursing and Personal Care Facilities (By Blocs and Areas)
Changes of life style How are ageing and birth ratio by areas evolving? ■国土のモニタリング Ratio of elderly population who are living alone, disabled, and cared in nursing and personal care facilities (by blocs and areas). A high percentage of elderly persons in the Kinki, Hokkaido, and Kyushu geographic regions live alone, a high percentage of those in the Chugoku and Shikoku geographic regions require daily care, and in the rural economic sphere, especially Okinawa, a high percentage are now housed in one of the three kinds of public long-term care facilities that accept long-term care insurance. (1) Elderly persons living alone (2) Elderly persons requiring daily care (percent of population aged 65 or over) (percent of population aged 65 or over) (2001year) (2000year) Nationwide Nationwide Kinki Chugoku Hokkaido Shikoku Kyusyu Kyushu Shikoku Chubu Okinawa Kinki Cyugoku Touhoku Kanto Hokkaido Cyuubu Kanto Hokuriku Hokuriku Touhoku Okinawa the three major urban economic spheres the three major urban economic spheres Tokyo urban economic spheres Tokyo urban economic spheres Nagoya urban economic spheres Nagoya urban economic spheres Kansai urban economic spheres Kansai urban economic spheres the rural economic spheres the rural economic spheres 0 2 4 6 8 101214161820 0 5 10 15 20 25 30 (%) (%) (3) Persons now housed in one of the three kinds of public long-term care facilities (2001year) (percent of population aged 65 or over) Nationwide Okinawa Note: (1)Percentage of elderly persons living alone was calculated by dividing the Hokuriku population aged 65 or older and living alone by total population Shikoku aged 65 or older Hokkaido (2) Does not include elderly housed in long-term care facilities or hospitalized. -

Tokushima-Komatsushima Port Tourist Information
Tokushima-Komatsushima Port Tourist Information http://www.mlit.go.jp/kankocho/cruise/ Tokushima Local Gourmet 【① Tokushima Ramen】This local ramen features a brown pork bone and soy sauce soup, sukiyaki-style sweet and spicy pork ribs, and is topped with a raw egg.【②Fish Katsu】Minced fish is mixed with curry powder, covered in breadcrumbs, and deep-fried to make this fish katsu. The spicy taste and fragrance are top notch. 【③ Mametentama】Mametentama is a type of okonomiyaki that is packed with sweetly cooked red kidney beans. The salty-sweet taste is addicting. 【④Handa Somen】These hand-pulled noodles are thicker than average noodles and are characterized by their chewiness. These refreshing noodles especially taste great when paired with sudachi. Location/View ① ② Access Season Year-round Discover TOKUSHIMA Japan ③ ④ Related links https://discovertokushima.net/en/culture/cuisines/ Contact Us【Tourism Policy Division Tokushima Prefectural Government】 TEL:+81-88-621-2337 / E-MAIL: [email protected]/ Website: https://discovertokushima.net/ Awa Indigo Dyed Textiles Indigo is also known as "Japan Blue" . Known for its high quality, production of Awa indigo flourished during the Edo period (1603-1867). In Tokushima, the tradition of Awa indigo dyed textiles continues to this day. Indigo dyed textiles are said to have antibacterial, insect repellent, and moisture retention properties. Aside from buying ready-made items, there are also workshops and facilities that allow you to make your own Awa indigo goods. Location/View Access Season Year-round Discover TOKUSHIMA Japan Related links https://discovertokushima.net/en/culture/crafts/awa-indigo- dyed/ Contact Us【Tourism Policy Division Tokushima Prefectural Government】 TEL:+81-88-621-2337 / E-MAIL: [email protected]/ Website: https://discovertokushima.net/ Arudeyo Tokushima (Tokushima Souvenir and Tourism Plaza) Located on the 1st floor of the Awa Odori Kaikan at the foot of Mt. -

Shikoku Food01.Pdf
Table of Contents Shikoku Island Kochi Prefecture 1 Brief Introduction of Shikoku 26-27 Brief Intoroduction of Kochi 28-56 Food Products and Companies in Kochi 28 Bellepoch Co., Ltd. Tokushima Prefecture 29 B.G.M. Kochi Co., Ltd. 2-3 Brief Intoroduction of Tokushima 30 Café du glace 4-13 Food Products and Companies in Tokushima 31 Fukutatsu Ltd. 4 Kurio Shoten Co., Ltd. 32 Graziemille Co., Ltd. 5 Hikari Foods Co., Ltd. 33 Happy-Farm Co., Ltd 6 Honke Matsuura Brewery Co., Ltd. 34 Hayashi Katsuobushiten Co., Ltd. 7 Naruto Kintoki Distillery 35 Ikeichi Saikaen Co., Ltd. 8 Nissin Shurui Co., Ltd. 36 Izuri Konbu Kaisan Co., Ltd. 9 Shoei Co., Ltd. 37 Katsuma Japan Co., Ltd. 10 Torishin Inc. 38 Kameizumi Sake Brewing Co., Ltd. 11 Yachiyo Ltd. 39 Kensho Co., Ltd. 12 Yaosugi Co., Ltd. 40 Kitagawa Village Yuzu Kingdom Co., Ltd. 13 Yoshidaya Ltd. 41 Kochi Ice Co., Ltd. 42 Kochi Brewery Inc. Kagawa Prefecture 43 Marine Gold Co., Ltd. 14-15 Brief Introduction of Kagawa 44 Moriki Suikoen Co., Ltd. 16-18 Food Products and Companies in Kagawa 45 Agricultural Production Corporation Noyuen 16 Kimura Kaisan Co., Ltd. 46 Nomura Irimame Kakoten Co., Ltd. 17 Somenya 47 Okabayashi Noen Co., Ltd. 18 Yamaroku Shouyu Co., Ltd. 48 Hori Okazaki Noen Co., Ltd. 19 Festivals in Shikoku 49 Sakura-do Japan 50 Shimantoseisan Limited Company Ehime Prefecture 51 Sukumo Shuzo Co., Ltd. 20-21 Biref Intoroduction of Ehime 52 Tanakashoku Co., Ltd. 22-24 Food Products and Companies in Ehime 53 Tokuyayoshi Shoji Co., Ltd.