To the Southern Hemisphere, Where They Have Long Been Pointed out As
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Darwin's Route from Ushuaia
Darwin’s Route from Ushuaia Departing from Ushuaia, retrace the route of Charles Darwin aboard HMS Beagle on an expedition cruise through the secluded Fuegian Archipelago at the bottom of South America. Our adventurous nine-day (eight-night) itinerary includes legendary Cape Horn and historic Wulaia Bay, as well as Glacier Alley, the penguin boisterous colonies on Tuckers and Magdalena islands, as well as the spectacular fjords that harbor Pía and Águila glaciers. While visiting Patagonia you'll also encounter massive ice fields, lush sub-polar forests and secluded beaches on islands that remain refreshingly remote and barely touched by civilization, a rare glimpse of what planet Earth must have been like before mankind. Midway through the journey, a half- day port call in Punta Arenas leaves plenty of time to explore a city rich in history, architecture and Patagonian culture before resuming the journey back to Ushuaia. (B = Breakfast, L = Lunch, D = Dinner) NOTE: You can also start in Punta Arenas. Similar itinerary available on the Stella Australis. Operates between September & April. Please ask for more details. Day 1: Ushuaia Check in at 409 San Martín Ave. in downtown Ushuaia between 10:00 and 17:00 (10 AM-5 PM) on the day of your cruise departure. 2016-2017 Season: Board the M/V Stella Australis at 17:30 (5:30 PM). 2017-2018 Season: Board the M/V Stella Australis at 18:00 (6 PM). After a welcoming toast and introduction of captain and crew, the ship departs for one of the most remote corners of planet Earth. -

Patagonian Explorer
PATAGONIAN EXPLORER Explore Patagonia on an adventurous five-day, four-night journeys between Ushuaia (Argentina) to Punta Arenas (Chile) through some of planet’s most remote places and incredible scenery. Discover the wild beauty of Patagonia on an Australis cruise that showcases the region’s pristine landscapes, rich wildlife and fascinating human history aboard an expedition ship that brings an extraordinary level of comfort and service to the uttermost edge of the world. Leaving the twinkling lights of Ushuaia behind, the vessel calls on fabled spots like Cape Horn and Wulaia Bay before cruising down the Beagle Channel. Continuing through the maze of islands, we’ll visit a number of glaciers, frozen giants that guard the southern flank of the ITINERARY Fuegian Archipelago, before entering the legendary Strait of Day 1 - Ushuaia Magellan. Last stop is the Isla Magdalena and its boisterous Check in at 160 Juan Manuel de Rosas Street in downtown Patagonia penguin colony before docking at Punta Arenas. Ushuaia between 10:00 and 17:00 (10 AM-5 PM) on the day of your cruise departure. Board the M/V Ventus Australis at 18:00 (6 PM). After a welcoming toast and introduction of captain and crew, the ship departs for one of the most remote corners of planet Earth. During the night we traverse the Beagle Channel and cross from Argentina into Chilean territorial waters. The lights of Ushuaia disappear as we turn into the narrow Murray Channel between Navarino and Hoste islands. Day 2 - Cape Horn - Wulaia Bay Around the break of dawn, Stella Australis crosses Nassau Bay and enters the remote archipelago that comprises Cape Horn National Park. -

Day 1: Punta Arenas Check in at 1385 O'higgins Street (Arturo Prat Port) in Punta Arenas Between 13:00 and 17:00 (1-5 PM) on T
Day 1: Punta Arenas Check in at 1385 O’Higgins Street (Arturo Prat Port) in Punta Arenas between 13:00 and 17:00 (1-5 PM) on the day of your cruise departure. Board the M/V Stella Australis or M/V Via Australis at 18:00 (6 PM). After a welcoming cocktail reception hosted by the captain and his crew, the ship departs for one of the remotest corners of planet Earth. During the night we cross the Strait of Magellan and enter the labyrinth of channels that define the southern extreme of Patagonian. The twinkling lights of Punta Arenas gradually fade into the distance as we enter the Whiteside Canal between Darwin Island and Isla Grande de Tierra del Fuego. Day 2: Ainsworth Bay – Tuckers Islets By dawn the ship is sailing up Admiralty Sound, a spectacular offshoot of the Strait of Magellan that stretches nearly halfway across Tierra del Fuego. The snow-capped peaks of Karukinka Natural Park stretch along the north side of the sound, while the south shore is defined by the deep fjords and broad bays of Alberto de Agostini National Park. We go ashore at Ainsworth Bay, which harbours copious bird life and a colony of southern elephant seals which can sometimes be spotted from the Zodiacs. Two guided excursions are available: one is along the edge of a stream, peat bog and beaver habitat to a waterfall-and-moss-covered rock face tucked deep inside a pristine sub-polar forest; the other is a more strenuous hike along the crest of a glacial moraine. -

Cristina Calderón Collecting Mapi (Rushes) for Traditional Basketry
Langscape Magazine is an extension of the voice of Terralingua. It supports our mission by educating the minds and hearts about the importance and value of biocultural diversity. We aim to promote a paradigm shift by illustrating biocultural diversity through scientific and traditional knowledge, within an appealing sensory context of articles, stories and art. ABOUT THE COVER PHOTOS Front: Caleta Douglas, Navarino Island, Chile. Cristina Calderón collecting mapi (rushes) for traditional basketry. Photo: Oliver Vogel, 2015 Back: North Pindos, Greece The return of the shepherds and their flocks. Photo: Stamos Abatis, 2015 Terralingua thanks the Logan and Kalliopeia Langscape Magazine is a Foundations for their generous support. Terralingua Publication Editor: Luisa Maffi Editorial Assistant: Christine Arpita Graphic Design: Imagine That Graphics Learn about Terralingua www.terralingua.org Receive Langscape Magazine by subscribing or by purchasing single copies. Details at: www.terralinguaubuntu.org Learn about Langscape Magazine www.terralinguaubuntu.org/langscape/home.htm © Terralingua 2016 Cristina Cristina Zárraga CalderónCalderón Cristina and material breakdown of our ancestral Yamana culture Memories of [Ed.: Yamana is another name for the Yagan culture]. The struggle to survive, first in the face of the overwhelming My Yagan presence of alcohol, then under the thumb of the Chilean military authorities. Grandmother “When my mom died in Mejillones and I was left in Tierra del Fuego, Chile orphaned, I went to live with my grandfather and Granny Julia, Karpakolikipa…” was born in Róbalo, on the 24th of May. And they “I say, so tells me my aunt, that when I was born After losing her parents, my grandmother was left under during the night, there was a storm from the south. -

The Spirit of the Southern Wind
El Espíritu del Viento del Sur The Spirit of the Southern Wind Estrecho de Magallanes, Canal Beagle & Cabo de Hornos Fotografías: Luis Bertea Rojas Textos: Denis Chevallay BOOK SERIES THE STRAIT OF MAGELLAN EXPLORATION CRUISE TO THE REMOTE REGIONS OF TIERRA DEL FUEGO AND STRAIT OF MAGELLAN EXCLUSIVE PHOTO EXPEDITION AND NATURE CRUISE A BOARD THE M/V FORREST [email protected] EXPEDITION CRUISE PHOTOGRAPHY, CIENce & EDUCATION www.patagoniaphotosafaris.com The Spirit of the Southern Wind El Espíritu del Viento del Sur Strait of Magellan, Beagle Channel & Cape Horn Photographs by Luis Bertea Rojas - Text by Denis Chevallay Glaciar en Seno Ballena-Isla Santa Ines Glacier on Whalesound-Santa Ines island Fuerte, bravío, impredecible e inclemente. Así es "El espíritu del viento del sur". Un viento que por siglos ha puesto a prueba a innumerables aventureros. Quienes fueron capaces de enfrentarlo, pudieron conocer parte de los misterios que aun encierra este hermoso rincón del planeta. La mayoría de los que embarcaron desde tierras lejanas no lo consiguieron, sin embargo, los vencedores tuvieron la oportunidad de dar a conocer al resto del mundo la belleza inhóspita de estos parajes y el coraje de los habitantes del llamado "fin del Mundo". Strong, fierce, unpredictable and inclement. These are just some words to describe “the spirit of the southern wind”. For centuries this wind has put innumerable adventurers to the test and only those capable of confronting the wind were able to learn a little about the mysteries that are still hidden in this beautiful corner of the planet. The majority of those explorers who set off from distant lands were unsuccessful in their attempts. -

Populations of a New Morphotype of Corrugate Lessonia Bory in the Beagle Channel, Sub-Antarctic Magellanic Ecoregion: a Possible Case of On-Going Speciation
cryptogamie Algologie 2020 ● 41 ● 11 DIRECTEUR DE LA PUBLICATION / PUBLICATION DIRECTOR : Bruno DAVID Président du Muséum national d’Histoire naturelle RÉDACTRICE EN CHEF / EDITOR-IN-CHIEF : Line LE GALL Muséum national d’Histoire naturelle ASSISTANTE DE RÉDACTION / ASSISTANT EDITOR : Audrina NEVEU ([email protected]) MISE EN PAGE / PAGE LAYOUT : Audrina NEVEU RÉDACTEURS ASSOCIÉS / ASSOCIATE EDITORS Ecoevolutionary dynamics of algae in a changing world Stacy KRUEGER-HADFIELD Department of Biology, University of Alabama, 1300 University Blvd, Birmingham, AL 35294 (United States) Jana KULICHOVA Department of Botany, Charles University, Prague (Czech Repubwlic) Cecilia TOTTI Dipartimento di Scienze della Vita e dell’Ambiente, Università Politecnica delle Marche, Via Brecce Bianche, 60131 Ancona (Italy) Phylogenetic systematics, species delimitation & genetics of speciation Sylvain FAUGERON UMI3614 Evolutionary Biology and Ecology of Algae, Departamento de Ecología, Facultad de Ciencias Biologicas, Pontificia Universidad Catolica de Chile, Av. Bernardo O’Higgins 340, Santiago (Chile) Marie-Laure GUILLEMIN Instituto de Ciencias Ambientales y Evolutivas, Universidad Austral de Chile, Valdivia (Chile) Diana SARNO Department of Integrative Marine Ecology, Stazione Zoologica Anton Dohrn, Villa Comunale, 80121 Napoli (Italy) Comparative evolutionary genomics of algae Nicolas BLOUIN Department of Molecular Biology, University of Wyoming, Dept. 3944, 1000 E University Ave, Laramie, WY 82071 (United States) Heroen VERBRUGGEN School of BioSciences, -
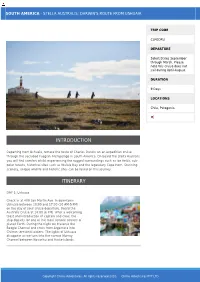
Introduction Itinerary
SOUTH AMERICA - STELLA AUSTRALIS: DARWIN'S ROUTE FROM USHUAIA TRIP CODE CLFODRU DEPARTURE Select Dates September through March. Please note this cruise does not sail during April-August. DURATION 9 Days LOCATIONS Chile, Patagonia INTRODUCTION Departing from Ushuaia, retrace the route of Charles Darwin on an expedition cruise through the secluded Fuegian Archipelago in South America. On board the Stella Australis you will find comfort whilst experiencing the rugged surroundings such as ice fields, sub- polar forests, historical sites such as Wulaia Bay and the legendary Cape Horn. Stunning scenery, unique wildlife and historic sites can be found on this journey. ITINERARY DAY 1: Ushuaia Check in at 409 San Martín Ave. in downtown Ushuaia between 10:00 and 17:00 (10 AM-5 PM) on the day of your cruise departure. Board the Australis Cruise at 18:00 (6 PM). After a welcoming toast and introduction of captain and crew, the ship departs for one of the most remote corners of planet Earth. During the night we traverse the Beagle Channel and cross from Argentina into Chilean territorial waters. The lights of Ushuaia disappear as we turn into the narrow Murray Channel between Navarino and Hoste islands. Copyright Chimu Adventures. All rights reserved 2020. Chimu Adventures PTY LTD SOUTH AMERICA - STELLA AUSTRALIS: DARWIN'S ROUTE FROM USHUAIA TRIP CODE CLFODRU DAY 2: Cape Horn - Wulaia Bay DEPARTURE Around the break of dawn, the Australis cruise crosses Nassau Bay and enters the remote archipelago that compromises Cape Horn National Select Dates September Park. Weather and sea conditions permitting, we through March. -

Tierra Del Fuego and Magellan Strait Contacts with Natives Reported in EnglishLanguage Newspapers (18511900) (Relevant Text Is Shown in Bold Type)
«Of Sailors and Savages» : Tierra del Fuego and Magellan Strait Contacts with natives reported in Englishlanguage newspapers (18511900) (relevant text is shown in bold type) PIONEER + SPEEDWELL [1851] Lyttelton Times (NZ), 18 September 1852 A PARTY OF MISSIONARIES STARVED TO DEATH. We have to narrate one of the most appalling stories that has ever appeared in a public journal. An English officer, Captain Gardiner, of the Royal Navy, who was sent out with six companions by the "Patagonian Missionary Society" to the neighbourhood of Cape Horn, has been literally starved to death — he, and his companions with him. The party consisted of Captain Allen Gardiner, R.N., superintendent; Mr. Williams, surgeon and catechist; Mr Maidment, catechist; John Irwin, carpenter; and John Badcock, John Bryant, & Pearce, Cornish fishermen. They left England in September, 1850, in the Ocean Queen; it having been promised that stores should be forwarded to them in June via the Falkland Islands; should they be unable to maintain their position at Picton Island, Beagle Channel, it was supposed that being provided with partially decked launches, they would fall back on Staten Island. The illfated party landed on Picton Island towards the conclusion of the year 1850. From the first they seem to have been annoyed in some measure by the natives, and to have been hunted backwards and forwards from the little island to what may be called the mainland of TerradelFuego. About the middle of April, 1851, Captain Gardiner begins to record in his diary, which, has been preserved to us, that they have provisions enough to last for two months, but some are very low. -

Journal of the Lagoda (Bark) of New Bedford, Mass., Mastered by Edward D
Journal of the Lagoda (Bark) of New Bedford, Mass., mastered by Edward D. Lewis, kept by Wilhelm Kromer, on voyage 11 April 1882–3 June 1886 Peter J. Collins, Volunteer [email protected] Mark Procknik, Librarian [email protected] 2 September 2014 Revised 23 December 2014 ii To my father’s memory Contents Introduction v March . 28 April . 32 1 1881 1 May................. 1 A ODHS 972B 33 August . 1 April 1882 . 33 September . 2 December 1882 . 33 November . 2 February 1883 . 33 2 1882 5 B ODHS 972C 35 June . 5 December 1883 . 35 July . 7 June 1884 . 35 August . 8 C ODHS 972D 37 September . 9 December 1883 . 37 October . 10 November . 12 D ODHS 972E 39 December . 15 December 1885 . 39 February 1886 . 39 3 1883 21 January . 21 E ODHS 972F 41 February . 25 Undated . 41 Annotations An aster*sk denotes an illegible letter or sequence of letters. Probable letter choices {ar,we}re enclosed in curly brackets. Square brackets clarify idiosyncratic [characteristic] words, and fill [in] those missing. Modern grammar conventions are used except with proper nouns. Emendations are documented in footnotes. iii iv CONTENTS Introduction Wilhelm Kromer was 22 when he joined the Bark Lagoda’s twelfth voyage out of New Bedford. This journey lasted from April 1882–June 1886 and took place in the Tierra del Fuego at the Chile–Argentina–Antarctica border. His series of six logbooks are the only known journals from the vessel under Capt. Edward D. Lewis. The first book is in mixed English, German, and French. The other five are in English with New Year’s and birthday blessings in German. -
Caleta Wulaia Reserva De La Biósfera
Ruta Patrimonial Nº16 Caleta Wulaia Reserva de la Biósfera Región de Magallanes y de la Antártica Chilena tapa.FH11 Tue Dec 04 16:02:54 2007 Page 2 C M Y CM MY CY CMY K Ruta Patrimonial CabCab Santiago 80º 05 26º 18 79º 51 26º 27 105º 28 109º 20 68º 44 26º 27 56º 30 33º 37 53º 46 PUERTO NAVARINO PUERTO LAGUNA WILLIAMS ROJAS ISLA C. WULAIA LAGO BUTTON NAVARINO LAGO WINDHOND RIO PTO. TORO DOUGLAS BAHIA WINDHOND PENINSULA SEÑORET Composite tapa.FH11 Tue Dec 04 16:02:54 2007 Page 3 C M Y CM MY CY CMY K abobo dede Hornos:Hornos: WulaiaWulaia Composite tapa.FH11 Tue Dec 04 16:02:54 2007 Page 4 C M Y CM MY CY CMY K Ubicación y accesos Location and access ways Ubicación Location La Ruta Patrimonial Cabo de Hornos-Wulaia The Heritage Road Cape Horn - Wulaia se ubica en la isla de Navarino, comuna Cabo is located in Navarino Island, district de Hornos, provincia Antártica perteneciente of Cape Horn, Antarctic Province a la XIIª Región de Magallanes. Se extiende pertaining to Region XII of en el extremo noroeste de la isla, en la ribera Magallanes. It is located in the north- sur del canal Beagle a 10 millas náuticas de east end of the island, in Beagle Usuahía y 55 kilómetros de Puerto Williams, Channel's south bank, 10 nautical miles capital comunal y principal núcleo urbano from Usuahia, and 55 kilometers from de la costa sur de Tierra del Fuego. Puerto Williams, the district capital and main urban community of Tierra Accesos del Fuego's south coast. -

Yaghan's, Explorers and Settlers
YAGHAN’S, EXPLORERS AND SETTLERS: 10,000 years of Southern Tierra del Fuego Archipelago History The Museum Permanent Exhibit Script Martin Gusinde Anthropological Museum · Puerto Williams - Chile Participants in Initiation Rite in the year 1922. (Martin Gusinde, last row, fourth position from left to right). Anthropos Institut. Sankt Augustin, Germany - Authorized Digital Copying Martin Gusinde Anthropological Museum Introduction INTROUDUCTION The creation of a museum on Navarino Island was an ambitious project that grew out of a deep interest in and concern for the island’s natural and cultural heritage. The initiative was originally proposed by the Chilean Navy, the same institution that built the Martín Gusinde Anthropological Museum (MAMG), which opened its doors in Puerto Williams in 1975. In the 1960s, before the museum was founded, a collection was begun of archeological material from the island’s coastal areas, along with some objects of historical interest from the first occupation by pioneers. This collection was exhibited in the now defunct Mixed School Nº in Puerto Williams. The collection was moved when the Martin Gusinde Museum was established, becoming part of that institution permanent collection. The Museum was named after the Austrian anthropologist and priest Martin Gusinde S.V.D. (1886-1969), who worked among the Yaghan and Selk’nam people from 1918 to 1924. His body of work offers the greatest collection of ethnographic studies about a world that was already on the verge of disappearing. His seminal work “The Indians of Tierra del Fuego” was published between 190 and 1974, and will forever remain the principal source of information we possess about the native people of Tierra del Fuego. -

Charles Darwin Ben Jonson Voyage of the Beagle Ii Love Freed from Ignor
PEOPLE MENTIONED IN WALDEN THERE WAS SOME RISK OF CHARLES ROBERT DARWIN’S 1 TURNING OUT AN IDLE MAN “NARRATIVE HISTORY” AMOUNTS TO FABULATION, THE REAL STUFF BEING MERE CHRONOLOGY 1. In the year that the Beagle sailed, Darwin was regarded as a budding geologist. His geology mentor, Woodwardian Professor of Geology at Cambridge Adam Sedgwick, would write while this young protégé was sailing around the world that: [He] is doing admirable work in South America, and has already sent home a collection above all price.... There was some risk of his turning out an idle man, but his character will now be fixed, and if God spares his life he will have a great name among the naturalists of Europe. HDT WHAT? INDEX THE PEOPLE OF WALDEN: CHARLES ROBERT DARWIN PEOPLE MENTIONED IN WALDEN “WALKING”: A tanned skin is something more than respectable, and perhaps olive is a fitter color than white for a man — a denizen of the woods. “The pale white man!” I do not wonder that the African pitied him. Darwin the naturalist says “A white man bathing by the side of a Tahitian was like a plant bleached by the gardener’s art compared with a fine, dark green one growing vigorously in the open fields.” Ben Jonson exclaims,— “How near to good is what is fair!” So I would say— How near to good is what is wild! Life consists with Wildness. The most alive is the wildest. Not yet subdued to man, its presence refreshes him. One who pressed forward incessantly and never rested from his labors, who grew fast and made infinite demands on life, would always find himself in a new country or wilderness, and surrounded by the raw material of life.