Fish Movement Through Intermittent Stream Channels
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-
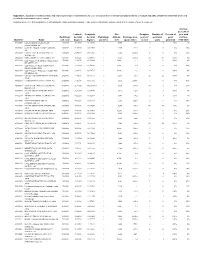
USGS Open-File Report 2009-1269, Appendix 1
Appendix 1. Summary of location, basin, and hydrological-regime characteristics for U.S. Geological Survey streamflow-gaging stations in Arizona and parts of adjacent states that were used to calibrate hydrological-regime models [Hydrologic provinces: 1, Plateau Uplands; 2, Central Highlands; 3, Basin and Range Lowlands; e, value not present in database and was estimated for the purpose of model development] Average percent of Latitude, Longitude, Site Complete Number of Percent of year with Hydrologic decimal decimal Hydrologic altitude, Drainage area, years of perennial years no flow, Identifier Name unit code degrees degrees province feet square miles record years perennial 1950-2005 09379050 LUKACHUKAI CREEK NEAR 14080204 36.47750 109.35010 1 5,750 160e 5 1 20% 2% LUKACHUKAI, AZ 09379180 LAGUNA CREEK AT DENNEHOTSO, 14080204 36.85389 109.84595 1 4,985 414.0 9 0 0% 39% AZ 09379200 CHINLE CREEK NEAR MEXICAN 14080204 36.94389 109.71067 1 4,720 3,650.0 41 0 0% 15% WATER, AZ 09382000 PARIA RIVER AT LEES FERRY, AZ 14070007 36.87221 111.59461 1 3,124 1,410.0 56 56 100% 0% 09383200 LEE VALLEY CR AB LEE VALLEY RES 15020001 33.94172 109.50204 1 9,440e 1.3 6 6 100% 0% NR GREER, AZ. 09383220 LEE VALLEY CREEK TRIBUTARY 15020001 33.93894 109.50204 1 9,440e 0.5 6 0 0% 49% NEAR GREER, ARIZ. 09383250 LEE VALLEY CR BL LEE VALLEY RES 15020001 33.94172 109.49787 1 9,400e 1.9 6 6 100% 0% NR GREER, AZ. 09383400 LITTLE COLORADO RIVER AT GREER, 15020001 34.01671 109.45731 1 8,283 29.1 22 22 100% 0% ARIZ. -

Roundtail Chub Repatriated to the Blue River
Volume 1 | Issue 2 | Summer 2015 Roundtail Chub Repatriated to the Blue River Inside this issue: With a fish exclusion barrier in place and a marked decline of catfish, the time was #TRENDINGNOW ................. 2 right for stocking Roundtail Chub into a remote eastern Arizona stream. New Initiative Launched for Southwest Native Trout.......... 2 On April 30, 2015, the Reclamation, and Marsh and Blue River. A total of 222 AZ 6-Species Conservation Department stocked 876 Associates LLC embarked on a Roundtail Chub were Agreement Renewal .............. 2 juvenile Roundtail Chub from mission to find, collect and stocked into the Blue River. IN THE FIELD ........................ 3 ARCC into the Blue River near bring into captivity some During annual monitoring, Recent and Upcoming AZGFD- the Juan Miller Crossing. Roundtail Chub for captive led Activities ........................... 3 five months later, Additional augmentation propagation from the nearest- Department staff captured Spikedace Stocked into Spring stockings to enhance the genetic neighbor population in Eagle Creek ..................................... 3 42 of the stocked chub, representation of the Blue River Creek. The Aquatic Research some of which had travelled BACK AT THE PONDS .......... 4 Roundtail Chub will be and Conservation Center as far as seven miles Native Fish Identification performed later this year. (ARCC) held and raised the upstream from the stocking Workshop at ARCC................ 4 offspring of those chub for Stockings will continue for the location. future stocking into the Blue next several years until that River. population is established in the Department biologists conducted annual Blue River and genetically In 2012, the partners delivered monitoring in subsequent mimics the wild source captive-raised juvenile years, capturing three chub population. -

Environmental Flows and Water Demands in Arizona
Environmental Flows and Water A University of Arizona Water Resources Research Center Project Demands in Arizona ater is an increasingly scarce resource and is essential for Arizona’s future. Figure 1. Elements of Environmental Flow WWith Arizona’s population growth and Occurring in Seasonal Hydrographs continued drought, citizens and water managers have been taking a closer look at water supplies in the state. Municipal, industrial, and agricul- tural water users are well-represented demand sectors, but water supplies and management to benefit the environment are not often consid- ered. This bulletin explains environmental water demands in Arizona and introduces information essential for considering environmental water demands in water management discussions. Considering water for the environment is impor- tant because humans have an interconnected and interdependent relationship with the envi- ronment. Nature provides us recreation oppor- tunities, economic benefits, and water supplies Data Source: to sustain our communities. USGS stream gage data Figure 2: Human Demand and Current Flow in Arizona Environmental water demands (or environmental flow) (circle size indicates relative amount of water) refers to how much water is needed in a watercourse to sustain a healthy ecosystem. Defining environmental water demand goes beyond the ecology and hydrol- Maximum ogy of a system and should include consideration for Flows how much water is required to achieve an agreed Industrial 40.8 maf Industrial SW Municipal upon level of river health, as determined by the GW 1% GW 8% water-using community. Arizona’s native ani- 4% mals and plants depend upon dynamic flows commonly described according to the natural Municipal SW flow regime. -

Cienegas Vanishing Climax Communities of the American
Hendrickson and Minckley Cienegas of the American Southwest 131 Abstract Cienegas The term cienega is here applied to mid-elevation (1,000-2,000 m) wetlands characterized by permanently saturated, highly organic, reducing soils. A depauperate Vanishing Climax flora dominated by low sedges highly adapted to such soils characterizes these habitats. Progression to cienega is Communities of the dependent on a complex association of factors most likely found in headwater areas. Once achieved, the community American Southwest appears stable and persistent since paleoecological data indicate long periods of cienega conditions, with infre- quent cycles of incision. We hypothesize the cienega to be an aquatic climax community. Cienegas and other marsh- land habitats have decreased greatly in Arizona in the Dean A. Hendrickson past century. Cultural impacts have been diverse and not Department of Zoology, well documented. While factors such as grazing and Arizona State University streambed modifications contributed to their destruction, the role of climate must also be considered. Cienega con- and ditions could be restored at historic sites by provision of ' constant water supply and amelioration of catastrophic W. L. Minckley flooding events. U.S. Fish and Wildlife Service Dexter Fish Hatchery Introduction and Department of Zoology Written accounts and photographs of early explorers Arizona State University and settlers (e.g., Hastings and Turner, 1965) indicate that most pre-1890 aquatic habitats in southeastern Arizona were different from what they are today. Sandy, barren streambeds (Interior Strands of Minckley and Brown, 1982) now lie entrenched between vertical walls many meters below dry valley surfaces. These same streams prior to 1880 coursed unincised across alluvial fills in shallow, braided channels, often through lush marshes. -

Riparian Buffer
Riparian Buffer A Technical Primer on Assessing Potential Water Quality Improvements for the Quantifying Benefits of Miyun Reservoir in Beijing, China Watershed Interventions September 2016 Prepared by: “Andrew” Feng Fang, Ph.D. Work supported by: Mark S. Kieser 536 E. Michigan Ave, Suite 300 Kalamazoo, MI 49007 USA The China Mega-City Water Fund (CMWF) was launched in August of 2015 in cooperation with the Beijing Forestry Society (BFS), China Biodiversity Conservation and Green Development Foundation (CBCGDF), International Union for Conservation of Nature (IUCN), and Forest Trends. Once fully operating, the CMWF will identify, fund, and help implement watershed improvement projects (“interventions”) to benefit water quality and water quantity in the Miyun Reservoir. The City of Beijing relies in part on this reservoir as a critical drinking water supply. A notable challenge facing the CMWF, and other water funds throughout the world, is the ability to reasonably estimate water quality and/or water quantity benefits associated with specific interventions. Recent 2016 efforts have established an operating framework for the CMWF that will evaluate various watershed interventions in the context of relatively simple, established performance metrics for water quality and water quantity benefits. Coupled with projected costs for such interventions, the CMWF will be able to assess, compare and optimize benefits associated with its investments in watershed improvements with this framework. This Technical Primer1 represents an initial examination of quantification methods that the CMWF and others may use to reliably estimate water resource benefits derived from particular land management interventions in the watershed of the Miyun Reservoir. This approach relies upon existing studies from both the Miyun Reservoir watershed and other basins in China. -

Gila Topminnow Revised Recovery Plan December 1998
GILA TOPMINNOW, Poeciliopsis occidentalis occidentalis, REVISED RECOVERY PLAN (Original Approval: March 15, 1984) Prepared by David A. Weedman Arizona Game and Fish Department Phoenix, Arizona for Region 2 U.S. Fish and Wildlife Service Albuquerque, New Mexico December 1998 Approved: Regional Director, U.S. Fish and Wildlife Service Date: Gila Topminnow Revised Recovery Plan December 1998 DISCLAIMER Recovery plans delineate reasonable actions required to recover and protect the species. The U.S. Fish and Wildlife Service (Service) prepares the plans, sometimes with the assistance of recovery teams, contractors, State and Federal Agencies, and others. Objectives are attained and any necessary funds made available subject to budgetary and other constraints affecting the parties involved, as well as the need to address other priorities. Time and costs provided for individual tasks are estimates only, and not to be taken as actual or budgeted expenditures. Recovery plans do not necessarily represent the views nor official positions or approval of any persons or agencies involved in the plan formulation, other than the Service. They represent the official position of the Service only after they have been signed by the Regional Director or Director as approved. Approved recovery plans are subject to modification as dictated by new findings, changes in species status, and the completion of recovery tasks. ii Gila Topminnow Revised Recovery Plan December 1998 ACKNOWLEDGMENTS Original preparation of the revised Gila topminnow Recovery Plan (1994) was done by Francisco J. Abarca 1, Brian E. Bagley, Dean A. Hendrickson 1 and Jeffrey R. Simms 1. That document was modified to this current version and the work conducted by those individuals is greatly appreciated and now acknowledged. -

Water, Summer 2008
Restoring Connections Vol. 11 Issue 2 Summer 2008 Newsletter of the Sky Island Alliance In this issue: A River Runs Beneath It by Randy Serraglio 4 Time and the Aquifer: Models and Long-term Thinking Water… by Julia Fonseca 5 Street and Public Rights-of-Way: Community Corridors of Heat & Dehydration OR Green Belts of Coolness & Rehydration by Brad Lancaster 6 A New Path for Water Use by Melissa Lamberton 7 The Power of Water by Janice Przybyl 8 Our special pull-out section on Ciénegas Monitoring Water with Remote Cameras by Sergio Avila 9 Waste Water / Holy Water by Ken Lamberton 10 Coyote Wells by Julia Fonseca 12 Finding Water in the Desert by Gary Williams 12 H2Oly Stories by Doug Bland 13 Restaurant Review: The Adobe Café & Bakery by Mary Rakestraw 14 Volunteers Make It Happen Rio Saracachi at Rancho Agua Fria in Sonora. by Sarah Williams 16 From the Director’s Desk: Swimming Holes and Groundwater by Matt Skroch, Executive Director Rivers and springs have been used to our several decades, or centuries, the water table will agricultural advantage for 12,000 years here, once again seep upwards to ground level, and though unsustainable groundwater mining is a those low points on the landscape we call rivers relatively new phenomena. We’ve discovered will flow once again. other temporary ways around the problem of increasing water scarcity — billions of dollars Either choice will eventually lead nature back to spent to pump water uphill for 330 miles being better days. The difference being that one choice Few experiences compare to the exhilaration of one spectacular example. -

Native Fish Restoration in Redrock Canyon
U.S. Department of the Interior Bureau of Reclamation Final Environmental Assessment Phoenix Area Office NATIVE FISH RESTORATION IN REDROCK CANYON U.S. Department of Agriculture Forest Service Southwestern Region Coronado National Forest Santa Cruz County, Arizona June 2008 Bureau of Reclamation Finding of No Significant Impact U.S. Forest Service Finding of No Significant Impact Decision Notice INTRODUCTION In accordance with the National Environmental Policy Act of 1969 (Public Law 91-190, as amended), the Bureau of Reclamation (Reclamation), as the lead Federal agency, and the Forest Service, U.S. Fish and Wildlife Service (FWS), and Arizona Game and Fish Department (AGFD), as cooperating agencies, have issued the attached final environmental assessment (EA) to disclose the potential environmental impacts resulting from construction of a fish barrier, removal of nonnative fishes with the piscicide antimycin A and/or rotenone, and restoration of native fishes and amphibians in Redrock Canyon on the Coronado National Forest (CNF). The Proposed Action is intended to improve the recovery status of federally listed fish and amphibians (Gila chub, Gila topminnow, Chiricahua leopard frog, and Sonora tiger salamander) and maintain a healthy native fishery in Redrock Canyon consistent with the CNF Plan and ongoing Endangered Species Act (ESA), Section 7(a)(2), consultation between Reclamation and the FWS. BACKGROUND The Proposed Action is part of a larger program being implemented by Reclamation to construct a series of fish barriers within the Gila River Basin to prevent the invasion of nonnative fishes into high-priority streams occupied by imperiled native fishes. This program is mandated by a FWS biological opinion on impacts of Central Arizona Project (CAP) water transfers to the Gila River Basin (FWS 2008a). -

San Pedro River Study Area Wild and Scenic River Eligibility Report
Prepared by the USDI Department of the Interior Bureau of Land Management, Tucson Field Office May 2016 The BLM manages more than 245 million acres of public land, the most of any Federal agency. This land, known as the National System of Public Lands, is primarily located in 12 Western states, including Alaska. The BLM also administers 700 million acres of sub-surface mineral estate throughout the nation. The BLM's mission is to manage and conserve the public lands for the use and enjoyment of present and future generations under our mandate of multiple-use and sustained yield. Cover photo – San Pedro River near Charleston courtesy of BLM/Bob Wick 2 San Pedro River Wild and Scenic River Study Area Eligibility Report This document consolidates resource information about the San Pedro River and one of its key tributaries, the Babocomari River. The purpose of this document is to provide a basis for reassessing the eligibility and suitability for inclusion of the San Pedro River and related free- flowing conditions and outstandingly remarkable river values into the National Wild and Scenic River System. I. BACKGROUND: The San Pedro River Wild and Scenic River Study Area Eligibility Report describes the information considered by the Bureau of Land Management (BLM) in the eligibility, and tentative re-classification of the San Pedro River for suitability analysis in the San Pedro Riparian National Conservation Area (SPRNCA) Resource Management Plan (RMP). The SPRNCA was established by Public Law (P.L.) 100-696 on Nov. 18, 1988 to preserve, protect, and enhance the aquatic, wildlife, archaeological, paleontological, scientific, cultural, educational, and recreational resources in the conservation area. -
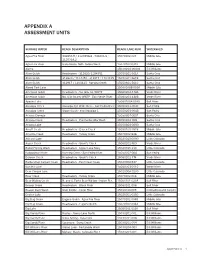
Appendix a Assessment Units
APPENDIX A ASSESSMENT UNITS SURFACE WATER REACH DESCRIPTION REACH/LAKE NUM WATERSHED Agua Fria River 341853.9 / 1120358.6 - 341804.8 / 15070102-023 Middle Gila 1120319.2 Agua Fria River State Route 169 - Yarber Wash 15070102-031B Middle Gila Alamo 15030204-0040A Bill Williams Alum Gulch Headwaters - 312820/1104351 15050301-561A Santa Cruz Alum Gulch 312820 / 1104351 - 312917 / 1104425 15050301-561B Santa Cruz Alum Gulch 312917 / 1104425 - Sonoita Creek 15050301-561C Santa Cruz Alvord Park Lake 15060106B-0050 Middle Gila American Gulch Headwaters - No. Gila Co. WWTP 15060203-448A Verde River American Gulch No. Gila County WWTP - East Verde River 15060203-448B Verde River Apache Lake 15060106A-0070 Salt River Aravaipa Creek Aravaipa Cyn Wilderness - San Pedro River 15050203-004C San Pedro Aravaipa Creek Stowe Gulch - end Aravaipa C 15050203-004B San Pedro Arivaca Cienega 15050304-0001 Santa Cruz Arivaca Creek Headwaters - Puertocito/Alta Wash 15050304-008 Santa Cruz Arivaca Lake 15050304-0080 Santa Cruz Arnett Creek Headwaters - Queen Creek 15050100-1818 Middle Gila Arrastra Creek Headwaters - Turkey Creek 15070102-848 Middle Gila Ashurst Lake 15020015-0090 Little Colorado Aspen Creek Headwaters - Granite Creek 15060202-769 Verde River Babbit Spring Wash Headwaters - Upper Lake Mary 15020015-210 Little Colorado Babocomari River Banning Creek - San Pedro River 15050202-004 San Pedro Bannon Creek Headwaters - Granite Creek 15060202-774 Verde River Barbershop Canyon Creek Headwaters - East Clear Creek 15020008-537 Little Colorado Bartlett Lake 15060203-0110 Verde River Bear Canyon Lake 15020008-0130 Little Colorado Bear Creek Headwaters - Turkey Creek 15070102-046 Middle Gila Bear Wallow Creek N. and S. Forks Bear Wallow - Indian Res. -

Proquest Dissertations
An ethnographic perspective on prehistoric platform mounds of the Tonto Basin, Central Arizona Item Type text; Dissertation-Reproduction (electronic) Authors Elson, Mark David, 1955- Publisher The University of Arizona. Rights Copyright © is held by the author. Digital access to this material is made possible by the University Libraries, University of Arizona. Further transmission, reproduction or presentation (such as public display or performance) of protected items is prohibited except with permission of the author. Download date 10/10/2021 00:20:59 Link to Item http://hdl.handle.net/10150/290644 INFORMATION TO USERS This manuscript has been reproduced from the microfihn master. UMI fihns the text directly from the orighud or copy submitted. Thus, some thesis and dissertation copies are in typewriter fiice, \^e others may be from any type of computer printer. The quality of this reproduction is dependent upon the quality of the copy submitted. Broken or indistinct print, colored or poor quality illustrations and photographs, print bleedthrough, substandard margins, and improper alignment can adversely affect reproduction. In the unlikely event that the author did not send UMI a complete manuscript and there are missing pages, these will be noted. Also, if unauthorized copyright material had to be removed, a note will indicate the deletion. Oversize materials (e.g., maps, drawings, charts) are reproduced by sectioning the original, b^inning at the upper left-hand comer and continuing from left to right in equal sections with small overlaps. Each original is also photographed in one exposure and is included in reduced form at the back of the book. -

James City County Perennial Stream Protocol Guidance Manual
James City County Perennial Stream Protocol Guidance Manual Purpose This protocol defines the procedure for making field determinations of the presence and the origin of a perennial stream. The procedure was developed to meet the requirement of James City County’s Chesapeake Bay Preservation Ordinance, Section 23-8, to perform site-specific field evaluations to determine the boundaries of Resource Protection Areas (RPA) and Section 23-10(2) to identify water bodies with perennial flow. Development of the protocol was finalized using funds from the Chesapeake Bay Implementation Grant #BAY-2007-14-PT awarded to the County by the Department of Conservation and Recreation (DCR). Introduction This protocol describes the process of determining whether a stream in James City County has perennial flow, and if necessary, determining the origin of the perennial stream. The origin of the perennial reach may occur as a transition from an ephemeral or intermittent reach or may occur at a spring with no upslope stream channel. The field identification of a perennial stream is based on the combination of hydrological, physical and biological characteristics of a stream. In the protocol, the field indicators of these characteristics are classified as primary or secondary and ranked using a weighted, four-tiered scoring system similar to the current system developed by the North Carolina Division of Water Quality (NCDWQ 2005) as modified to address conditions in James City County. A stream reach is classified as perennial based on the overall score but with consideration of other supporting information such as long term monitoring or historic information in certain situations.