Arxiv:1610.05765V3 [Astro-Ph.EP] 27 Feb 2017 3.3 Microlensing and Direct Imaging
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-
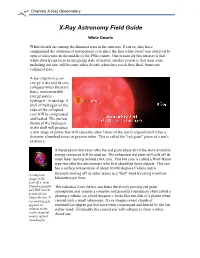
White Dwarfs
Chandra X-Ray Observatory X-Ray Astronomy Field Guide White Dwarfs White dwarfs are among the dimmest stars in the universe. Even so, they have commanded the attention of astronomers ever since the first white dwarf was observed by optical telescopes in the middle of the 19th century. One reason for this interest is that white dwarfs represent an intriguing state of matter; another reason is that most stars, including our sun, will become white dwarfs when they reach their final, burnt-out collapsed state. A star experiences an energy crisis and its core collapses when the star's basic, non-renewable energy source - hydrogen - is used up. A shell of hydrogen on the edge of the collapsed core will be compressed and heated. The nuclear fusion of the hydrogen in the shell will produce a new surge of power that will cause the outer layers of the star to expand until it has a diameter a hundred times its present value. This is called the "red giant" phase of a star's existence. A hundred million years after the red giant phase all of the star's available energy resources will be used up. The exhausted red giant will puff off its outer layer leaving behind a hot core. This hot core is called a Wolf-Rayet type star after the astronomers who first identified these objects. This star has a surface temperature of about 50,000 degrees Celsius and is A composite furiously boiling off its outer layers in a "fast" wind traveling 6 million image of the kilometers per hour. -

A Review of Possible Planetary Atmospheres in the TRAPPIST-1 System
Space Sci Rev (2020) 216:100 https://doi.org/10.1007/s11214-020-00719-1 A Review of Possible Planetary Atmospheres in the TRAPPIST-1 System Martin Turbet1 · Emeline Bolmont1 · Vincent Bourrier1 · Brice-Olivier Demory2 · Jérémy Leconte3 · James Owen4 · Eric T. Wolf5 Received: 14 January 2020 / Accepted: 4 July 2020 / Published online: 23 July 2020 © The Author(s) 2020 Abstract TRAPPIST-1 is a fantastic nearby (∼39.14 light years) planetary system made of at least seven transiting terrestrial-size, terrestrial-mass planets all receiving a moderate amount of irradiation. To date, this is the most observationally favourable system of po- tentially habitable planets known to exist. Since the announcement of the discovery of the TRAPPIST-1 planetary system in 2016, a growing number of techniques and approaches have been used and proposed to characterize its true nature. Here we have compiled a state- of-the-art overview of all the observational and theoretical constraints that have been ob- tained so far using these techniques and approaches. The goal is to get a better understanding of whether or not TRAPPIST-1 planets can have atmospheres, and if so, what they are made of. For this, we surveyed the literature on TRAPPIST-1 about topics as broad as irradiation environment, planet formation and migration, orbital stability, effects of tides and Transit Timing Variations, transit observations, stellar contamination, density measurements, and numerical climate and escape models. Each of these topics adds a brick to our understand- ing of the likely—or on the contrary unlikely—atmospheres of the seven known planets of the system. -

The Angular Momentum of Condensations Within Elephant Trunks
A&A 503, 477–482 (2009) Astronomy DOI: 10.1051/0004-6361/200912238 & c ESO 2009 Astrophysics The angular momentum of condensations within elephant trunks V. L ora 1,A.C.Raga2, and A. Esquivel2 1 Instituto de Astronomía, Universidad Nacional Autónoma de México, Ap. 70-468, 04510, D.F México 2 Instituto de Ciencias Nucleares, Universidad Nacional Autónoma de México, Ap. 70-543, 04510, D.F México e-mail: [email protected] Received 31 March 2009 / Accepted 22 May 2009 ABSTRACT Aims. The radiation from newly born stars photoevaporates their parental neutral cloud, leading to the formation of dense clumps that will eventually form stars. Methods. We present 3D simulations of the interaction of a neutral cloud with an external ionising radiation field, and compute the angular momenta of these collapsing clumps. Results. The angular momenta of these collapsing clumps show that they have preferential orientations mostly perpendicular to the direction of the incident ionising photon field. Therefore, the axes of the jet systems that will be eventually ejected (from the star+accretion disk systems that will form) will be oriented approximately perpendicular to the direction to the photoionising source. Key words. ISM: kinematics and dynamics – ISM: clouds – ISM: HII regions – stars: formation 1. Introduction of elephant trunks is quite small, their alignment approximately perpendicular to the direction to the ionising photon source The radiation from newly born stars photoionises and erodes the might be indicative of a systematic alignment. This alignment parental cloud, producing structures such as the so-called ele- implies that the angular momenta of the low mass star+disk sys- phant trunks. -

A Spitzer Survey of Protoplanetary Disk Dust in the Young Serpens Cloud: How Do Dust Characteristics Evolve with Time?
The Astrophysical Journal, 714:778–798, 2010 May 1 doi:10.1088/0004-637X/714/1/778 C 2010. The American Astronomical Society. All rights reserved. Printed in the U.S.A. A SPITZER SURVEY OF PROTOPLANETARY DISK DUST IN THE YOUNG SERPENS CLOUD: HOW DO DUST CHARACTERISTICS EVOLVE WITH TIME? Isa Oliveira1,2, Klaus M. Pontoppidan2, Bruno Mer´ın3, Ewine F. van Dishoeck1,4, Fred Lahuis1,5, Vincent C. Geers6, Jes K. Jørgensen7, Johan Olofsson8, Jean-Charles Augereau8, and Joanna M. Brown4 1 Leiden Observatory, Leiden University, P.O. Box 9513, 2300 RA Leiden, The Netherlands; [email protected] 2 California Institute of Technology, Division for Geological and Planetary Sciences, MS 150-21, Pasadena, CA 91125, USA 3 Herschel Science Center, European Space Agency (ESA), P.O. Box 78, 28691 Villanueva de la Canada˜ (Madrid), Spain 4 Max-Planck Institut fur¨ Extraterrestrische Physik, Giessenbachstrasse 1, 85748 Garching, Germany 5 SRON Netherlands Institute for Space Research, P.O. Box 800, 9700 AV Groningen, The Netherlands 6 University of Toronto, 50 St. George St., Toronto, ON M5R 2W9, Canada 7 Centre for Star and Planet Formation, Natural History Museum of Denmark, University of Copenhagen, Øster Voldgade 5-7, DK-1350 Copenhagen, Denmark 8 Laboratoire d’Astrophysique de Grenoble, Universite´ Joseph Fourier, CNRS, UMR 5571, Grenoble, France Received 2009 December 28; accepted 2010 March 16; published 2010 April 14 ABSTRACT We present Spitzer InfraRed Spectrograph (IRS) mid-infrared (5–35 μm) spectra of a complete flux-limited sample (3mJyat8μm) of young stellar object (YSO) candidates selected on the basis of their infrared colors in the Serpens Molecular Cloud. -

The Potential to Form Planets in the Orion Nebula ! Rita Mann Plaskett Fellow NRC-Herzberg
The Potential to Form Planets in the Orion Nebula ! Rita Mann Plaskett Fellow NRC-Herzberg ! James Di Francesco, Sean Andrews, Jonathan Williams, Doug Johnstone, John Bally, Meredith Hughes, Luca Ricci, Brenda Matthews Protoplanetary Disks in the Orion Nebula Most stars form in rich clusters Our Solar System formed in a massive star forming environment. To understand planet formation, we need to study disk properties in massive star forming regions! HST images of protostars in Orion Orion Nebula Cluster Trapezium Cluster • Thousands of protostars • Ages ~ 1-2 Myr • Distance ~ 400 pc 1 • θ Ori C, 40M¤, O6 SpT Orion Nebula Cluster Massive Stars • Hostile environment • Many low mass stars near ϑ1C have teardrop shaped morphologies Low Mass Stars PROPLYDS: PROtoPLanetarY DiskS Photoevaporating Proplyds -7 VLA mass-loss rate of 10 M¤/yr Churchwell et al. (1987) Mdisk < 0.1 M¤ Evaporation Timescales < 1 Myr Material removed too quickly! C.R. O’Dell Is planet formation inhibited in rich clusters? Disk Masses in Orion: Previous Attempts Millimeter Wavelength Interferometers (clustered disks) BIMA OVRO PdBI Mundy et al. (1995) Bally et al. (1998) Lada (1999) λ = 3.5 mm λ = 1.3 mm λ = 1.3 mm low sensitivity no detections never published Mdisk ≲ 15 MJUP Radio-Submillimeter SED -0.1 2-4 Ffree-free ~ ν + Fdust ~ ν λ = 1 cm Radio-Submillimeter SED -0.1 2-4 Ffree-free ~ ν + Fdust ~ ν λ = 1 cm 1 mm Radio-Submillimeter SED -0.1 2-4 Ffree-free ~ ν + Fdust ~ ν λ = 1 cm 1 mm Higher frequency observations: more sensitive to dust emission! Interferometry with -

The Sun, Yellow Dwarf Star at the Heart of the Solar System NASA.Gov, Adapted by Newsela Staff
Name: ______________________________ Period: ______ Date: _____________ Article of the Week Directions: Read the following article carefully and annotate. You need to include at least 1 annotation per paragraph. Be sure to include all of the following in your total annotations. Annotation = Marking the Text + A Note of Explanation 1. Great Idea or Point – Write why you think it is a good idea or point – ! 2. Confusing Point or Idea – Write a question to ask that might help you understand – ? 3. Unknown Word or Phrase – Circle the unknown word or phrase, then write what you think it might mean based on context clues or your word knowledge – 4. A Question You Have – Write a question you have about something in the text – ?? 5. Summary – In a few sentences, write a summary of the paragraph, section, or passage – # The sun, yellow dwarf star at the heart of the solar system NASA.gov, adapted by Newsela staff Picture and Caption ___________________________ ___________________________ ___________________________ Paragraph #1 ___________________________ ___________________________ This image shows an enormous eruption of solar material, called a coronal mass ejection, spreading out into space, captured by NASA's Solar Dynamics ___________________________ Observatory on January 8, 2002. Paragraph #2 Para #1 The sun is a hot ball made of glowing gases and is a type ___________________________ of star known as a yellow dwarf. It is at the heart of our solar system. ___________________________ Para #2 The solar system consists of everything that orbits the ___________________________ sun. The sun's gravity holds the solar system together, by keeping everything from planets to bits of dust in its orbit. -
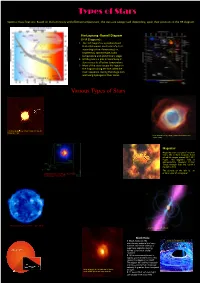
Brown Dwarf: White Dwarf: Hertzsprung -Russell Diagram (H-R
Types of Stars Spectral Classifications: Based on the luminosity and effective temperature , the stars are categorized depending upon their positions in the HR diagram. Hertzsprung -Russell Diagram (H-R Diagram) : 1. The H-R Diagram is a graphical tool that astronomers use to classify stars according to their luminosity (i.e. brightness), spectral type, color, temperature and evolutionary stage. 2. HR diagram is a plot of luminosity of stars versus its effective temperature. 3. Most of the stars occupy the region in the diagram along the line called the main sequence. During that stage stars are fusing hydrogen in their cores. Various Types of Stars Brown Dwarf: White Dwarf: Brown dwarfs are sub-stellar objects After a star like the sun exhausts its nuclear that are not massive enough to sustain fuel, it loses its outer layer as a "planetary nuclear fusion processes. nebula" and leaves behind the remnant "white Since, comparatively they are very cold dwarf" core. objects, it is difficult to detect them. Stars with initial masses Now there are ongoing efforts to study M < 8Msun will end as white dwarfs. them in infrared wavelengths. A typical white dwarf is about the size of the This picture shows a brown dwarf around Earth. a star HD3651 located 36Ly away in It is very dense and hot. A spoonful of white constellation of Pisces. dwarf material on Earth would weigh as much as First directly detected Brown Dwarf HD 3651B. few tons. Image by: ESO The image is of Helix nebula towards constellation of Aquarius hosts a White Dwarf Helix Nebula 6500Ly away. -

Supernovae Sparked by Dark Matter in White Dwarfs
Supernovae Sparked By Dark Matter in White Dwarfs Javier F. Acevedog and Joseph Bramanteg;y gThe Arthur B. McDonald Canadian Astroparticle Physics Research Institute, Department of Physics, Engineering Physics, and Astronomy, Queen's University, Kingston, Ontario, K7L 2S8, Canada yPerimeter Institute for Theoretical Physics, Waterloo, Ontario, N2L 2Y5, Canada November 27, 2019 Abstract It was recently demonstrated that asymmetric dark matter can ignite supernovae by collecting and collapsing inside lone sub-Chandrasekhar mass white dwarfs, and that this may be the cause of Type Ia supernovae. A ball of asymmetric dark matter accumulated inside a white dwarf and collapsing under its own weight, sheds enough gravitational potential energy through scattering with nuclei, to spark the fusion reactions that precede a Type Ia supernova explosion. In this article we elaborate on this mechanism and use it to place new bounds on interactions between nucleons 6 16 and asymmetric dark matter for masses mX = 10 − 10 GeV. Interestingly, we find that for dark matter more massive than 1011 GeV, Type Ia supernova ignition can proceed through the Hawking evaporation of a small black hole formed by the collapsed dark matter. We also identify how a cold white dwarf's Coulomb crystal structure substantially suppresses dark matter-nuclear scattering at low momentum transfers, which is crucial for calculating the time it takes dark matter to form a black hole. Higgs and vector portal dark matter models that ignite Type Ia supernovae are explored. arXiv:1904.11993v3 [hep-ph] 26 Nov 2019 Contents 1 Introduction 2 2 Dark matter capture, thermalization and collapse in white dwarfs 4 2.1 Dark matter capture . -

Chapter 16 the Sun and Stars
Chapter 16 The Sun and Stars Stargazing is an awe-inspiring way to enjoy the night sky, but humans can learn only so much about stars from our position on Earth. The Hubble Space Telescope is a school-bus-size telescope that orbits Earth every 97 minutes at an altitude of 353 miles and a speed of about 17,500 miles per hour. The Hubble Space Telescope (HST) transmits images and data from space to computers on Earth. In fact, HST sends enough data back to Earth each week to fill 3,600 feet of books on a shelf. Scientists store the data on special disks. In January 2006, HST captured images of the Orion Nebula, a huge area where stars are being formed. HST’s detailed images revealed over 3,000 stars that were never seen before. Information from the Hubble will help scientists understand more about how stars form. In this chapter, you will learn all about the star of our solar system, the sun, and about the characteristics of other stars. 1. Why do stars shine? 2. What kinds of stars are there? 3. How are stars formed, and do any other stars have planets? 16.1 The Sun and the Stars What are stars? Where did they come from? How long do they last? During most of the star - an enormous hot ball of gas day, we see only one star, the sun, which is 150 million kilometers away. On a clear held together by gravity which night, about 6,000 stars can be seen without a telescope. -

OLLI: the Birth, Life, and Death Of
The Birth, Life, and Death of Stars The Osher Lifelong Learning Institute Florida State University Jorge Piekarewicz Department of Physics [email protected] Schedule: September 29 – November 3 Time: 11:30am – 1:30pm Location: Pepper Center, Broad Auditorium J. Piekarewicz (FSU-Physics) The Birth, Life, and Death of Stars Fall 2014 1 / 12 Ten Compelling Questions What is the raw material for making stars and where did it come from? What forces of nature contribute to energy generation in stars? How and where did the chemical elements form? ? How long do stars live? How will our Sun die? How do massive stars explode? ? What are the remnants of such stellar explosions? What prevents all stars from dying as black holes? What is the minimum mass of a black hole? ? What is role of FSU researchers in answering these questions? J. Piekarewicz (FSU-Physics) The Birth, Life, and Death of Stars Fall 2014 2 / 12 The Birth of Carbon: The Triple-Alpha Reaction The A=5 and A=8 Bottle-Neck 5 −22 p + α ! Li ! p + α (t1=2 ≈10 s) 8 −16 α + α ! Be ! α + α (t1=2 ≈10 s) BBN does not generate any heavy elements! He-ashes fuse in the hot( T ≈108 K) and dense( n≈1028 cm−3) core 8 −8 Physics demands a tiny concentration of Be (n8=n4 ≈10 ) Carbon is formed: α + α ! 8Be + α ! 12C + γ (7:367 MeV) Every atom in our body has been formed in stellar cores! J. Piekarewicz (FSU-Physics) The Birth, Life, and Death of Stars Fall 2014 3 / 12 Stellar Nucleosynthesis: From Carbon to Iron Stars are incredibly efficient thermonuclear furnaces Heavier He-ashes fuse to produce: C,N,O,F,Ne,Na,Mg,.. -

Abstracts Connecting to the Boston University Network
20th Cambridge Workshop: Cool Stars, Stellar Systems, and the Sun July 29 - Aug 3, 2018 Boston / Cambridge, USA Abstracts Connecting to the Boston University Network 1. Select network ”BU Guest (unencrypted)” 2. Once connected, open a web browser and try to navigate to a website. You should be redirected to https://safeconnect.bu.edu:9443 for registration. If the page does not automatically redirect, go to bu.edu to be brought to the login page. 3. Enter the login information: Guest Username: CoolStars20 Password: CoolStars20 Click to accept the conditions then log in. ii Foreword Our story starts on January 31, 1980 when a small group of about 50 astronomers came to- gether, organized by Andrea Dupree, to discuss the results from the new high-energy satel- lites IUE and Einstein. Called “Cool Stars, Stellar Systems, and the Sun,” the meeting empha- sized the solar stellar connection and focused discussion on “several topics … in which the similarity is manifest: the structures of chromospheres and coronae, stellar activity, and the phenomena of mass loss,” according to the preface of the resulting, “Special Report of the Smithsonian Astrophysical Observatory.” We could easily have chosen the same topics for this meeting. Over the summer of 1980, the group met again in Bonas, France and then back in Cambridge in 1981. Nearly 40 years on, I am comfortable saying these workshops have evolved to be the premier conference series for cool star research. Cool Stars has been held largely biennially, alternating between North America and Europe. Over that time, the field of stellar astro- physics has been upended several times, first by results from Hubble, then ROSAT, then Keck and other large aperture ground-based adaptive optics telescopes. -

Monday, November 13, 2017 WHAT DOES IT MEAN to BE HABITABLE? 8:15 A.M. MHRGC Salons ABCD 8:15 A.M. Jang-Condell H. * Welcome C
Monday, November 13, 2017 WHAT DOES IT MEAN TO BE HABITABLE? 8:15 a.m. MHRGC Salons ABCD 8:15 a.m. Jang-Condell H. * Welcome Chair: Stephen Kane 8:30 a.m. Forget F. * Turbet M. Selsis F. Leconte J. Definition and Characterization of the Habitable Zone [#4057] We review the concept of habitable zone (HZ), why it is useful, and how to characterize it. The HZ could be nicknamed the “Hunting Zone” because its primary objective is now to help astronomers plan observations. This has interesting consequences. 9:00 a.m. Rushby A. J. Johnson M. Mills B. J. W. Watson A. J. Claire M. W. Long Term Planetary Habitability and the Carbonate-Silicate Cycle [#4026] We develop a coupled carbonate-silicate and stellar evolution model to investigate the effect of planet size on the operation of the long-term carbon cycle, and determine that larger planets are generally warmer for a given incident flux. 9:20 a.m. Dong C. F. * Huang Z. G. Jin M. Lingam M. Ma Y. J. Toth G. van der Holst B. Airapetian V. Cohen O. Gombosi T. Are “Habitable” Exoplanets Really Habitable? A Perspective from Atmospheric Loss [#4021] We will discuss the impact of exoplanetary space weather on the climate and habitability, which offers fresh insights concerning the habitability of exoplanets, especially those orbiting M-dwarfs, such as Proxima b and the TRAPPIST-1 system. 9:40 a.m. Fisher T. M. * Walker S. I. Desch S. J. Hartnett H. E. Glaser S. Limitations of Primary Productivity on “Aqua Planets:” Implications for Detectability [#4109] While ocean-covered planets have been considered a strong candidate for the search for life, the lack of surface weathering may lead to phosphorus scarcity and low primary productivity, making aqua planet biospheres difficult to detect.