Legionella Pneumophila
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

NCTC) Bacterial Strain Equivalents to American Type Culture Collection (ATCC) Bacterial Strains
This list shows National Collection of Type Cultures (NCTC) bacterial strain equivalents to American Type Culture Collection (ATCC) bacterial strains. NCTC Number CurrentName ATCC Number NCTC 7212 Acetobacter pasteurianus ATCC 23761 NCTC 10138 Acholeplasma axanthum ATCC 25176 NCTC 10171 Acholeplasma equifetale ATCC 29724 NCTC 10128 Acholeplasma granularum ATCC 19168 NCTC 10172 Acholeplasma hippikon ATCC 29725 NCTC 10116 Acholeplasma laidlawii ATCC 23206 NCTC 10134 Acholeplasma modicum ATCC 29102 NCTC 10188 Acholeplasma morum ATCC 33211 NCTC 10150 Acholeplasma oculi ATCC 27350 NCTC 10198 Acholeplasma parvum ATCC 29892 NCTC 8582 Achromobacter denitrificans ATCC 15173 NCTC 10309 Achromobacter metalcaligenes ATCC 17910 NCTC 10807 Achromobacter xylosoxidans subsp. xylosoxidans ATCC 27061 NCTC 10808 Achromobacter xylosoxidans subsp. xylosoxidans ATCC 17062 NCTC 10809 Achromobacter xylosoxidans subsp. xylosoxidans ATCC 27063 NCTC 12156 Acinetobacter baumannii ATCC 19606 NCTC 10303 Acinetobacter baumannii ATCC 17904 NCTC 7844 Acinetobacter calcoaceticus ATCC 15308 NCTC 12983 Acinetobacter calcoaceticus ATCC 23055 NCTC 8102 acinetobacter dna group 13 ATCC 17903 NCTC 10304 Acinetobacter genospecies 13 ATCC 17905 NCTC 10306 Acinetobacter haemolyticus ATCC 17907 NCTC 10305 Acinetobacter haemolyticus subsp haemolyticus ATCC 17906 NCTC 10308 Acinetobacter johnsonii ATCC 17909 NCTC 10307 Acinetobacter junii ATCC 17908 NCTC 5866 Acinetobacter lwoffii ATCC 15309 NCTC 12870 Actinobacillus delphinicola ATCC 700179 NCTC 8529 Actinobacillus equuli ATCC 19392 -

Legionella Shows a Diverse Secondary Metabolism Dependent on a Broad Spectrum Sfp-Type Phosphopantetheinyl Transferase
Legionella shows a diverse secondary metabolism dependent on a broad spectrum Sfp-type phosphopantetheinyl transferase Nicholas J. Tobias1, Tilman Ahrendt1, Ursula Schell2, Melissa Miltenberger1, Hubert Hilbi2,3 and Helge B. Bode1,4 1 Fachbereich Biowissenschaften, Merck Stiftungsprofessur fu¨r Molekulare Biotechnologie, Goethe Universita¨t, Frankfurt am Main, Germany 2 Max von Pettenkofer Institute, Ludwig-Maximilians-Universita¨tMu¨nchen, Munich, Germany 3 Institute of Medical Microbiology, University of Zu¨rich, Zu¨rich, Switzerland 4 Buchmann Institute for Molecular Life Sciences, Goethe Universita¨t, Frankfurt am Main, Germany ABSTRACT Several members of the genus Legionella cause Legionnaires’ disease, a potentially debilitating form of pneumonia. Studies frequently focus on the abundant number of virulence factors present in this genus. However, what is often overlooked is the role of secondary metabolites from Legionella. Following whole genome sequencing, we assembled and annotated the Legionella parisiensis DSM 19216 genome. Together with 14 other members of the Legionella, we performed comparative genomics and analysed the secondary metabolite potential of each strain. We found that Legionella contains a huge variety of biosynthetic gene clusters (BGCs) that are potentially making a significant number of novel natural products with undefined function. Surprisingly, only a single Sfp-like phosphopantetheinyl transferase is found in all Legionella strains analyzed that might be responsible for the activation of all carrier proteins in primary (fatty acid biosynthesis) and secondary metabolism (polyketide and non-ribosomal peptide synthesis). Using conserved active site motifs, we predict Submitted 29 June 2016 some novel compounds that are probably involved in cell-cell communication, Accepted 25 October 2016 Published 24 November 2016 differing to known communication systems. -

The Risk to Human Health from Free-Living Amoebae Interaction with Legionella in Drinking and Recycled Water Systems
THE RISK TO HUMAN HEALTH FROM FREE-LIVING AMOEBAE INTERACTION WITH LEGIONELLA IN DRINKING AND RECYCLED WATER SYSTEMS Dissertation submitted by JACQUELINE MARIE THOMAS BACHELOR OF SCIENCE (HONOURS) AND BACHELOR OF ARTS, UNSW In partial fulfillment of the requirements for the award of DOCTOR OF PHILOSOPHY in ENVIRONMENTAL ENGINEERING SCHOOL OF CIVIL AND ENVIRONMENTAL ENGINEERING FACULTY OF ENGINEERING MAY 2012 SUPERVISORS Professor Nicholas Ashbolt Office of Research and Development United States Environmental Protection Agency Cincinnati, Ohio USA and School of Civil and Environmental Engineering Faculty of Engineering The University of New South Wales Sydney, Australia Professor Richard Stuetz School of Civil and Environmental Engineering Faculty of Engineering The University of New South Wales Sydney, Australia Doctor Torsten Thomas School of Biotechnology and Biomolecular Sciences Faculty of Science The University of New South Wales Sydney, Australia ORIGINALITY STATEMENT '1 hereby declare that this submission is my own work and to the best of my knowledge it contains no materials previously published or written by another person, or substantial proportions of material which have been accepted for the award of any other degree or diploma at UNSW or any other educational institution, except where due acknowledgement is made in the thesis. Any contribution made to the research by others, with whom 1 have worked at UNSW or elsewhere, is explicitly acknowledged in the thesis. I also declare that the intellectual content of this thesis is the product of my own work, except to the extent that assistance from others in the project's design and conception or in style, presentation and linguistic expression is acknowledged.' Signed ~ ............................ -

Genetic and Functional Studies of the Mip Protein of Legionella
t1.ì. Genetic and Functional Studies of the Mip Protein of Legionella Rodney Mark Ratcliff, BSc (Hons)' MASM Infectious Diseases Laboratories Institute of Medical and Veterinary Science and Department of Microbiology and Immunology UniversitY of Adelaide. Adelaide, South Australia A thesis submitted to the University of Adelaide for the degree of I)octor of Philosophy 15'h March 2000 amended 14th June 2000 Colonies of several Legionella strains on charcoal yeast extract agar (CYE) after 4 days incubation at 37"C in air. Various magnifications show typical ground-glass opalescent appearance. Some pure strains exhibit pleomorphic growth or colour. The top two photographs demonstrate typical red (LH) and blue-white (RH) fluorescence exhibited by some species when illuminated by a Woods (IJV) Lamp. * t Table of Contents .1 Chapter One: Introduction .1 Background .............'. .2 Morphology and TaxonomY J Legionellosis ............. 5 Mode of transmission "..'....'. 7 Environmental habitat 8 Interactions between Legionella and phagocytic hosts 9 Attachment 11 Engulfment and internalisation.'.. 13 Intracellular processing 13 Intracellular replication of legionellae .. " "' " "' 15 Host cell death and bacterial release 18 Virulence (the Genetic factors involved with intracellular multiplication and host cell killing .20 icm/dot system) Legiolysin .25 Msp (Znn* metaloprotease) ...'..... .25 .28 Lipopolysaccharide .29 The association of flagella with disease.. .30 Type IV fimbriae.... .31 Major outer membrane proteins....'.......'. JJ Heat shock proteins'.'. .34 Macrophage infectivity potentiator (Mip) protein Virulenceiraits of Legionella species other than L. pneumophila..........' .39 phylogeny .41 Chapter One (continued): Introduction to bacterial classification and .41 Identificati on of Legionella...'.,..'.. .46 Phylogeny .52 Methods of phylogenetic analysis' .53 Parsimony methods.'.. .55 Distance methods UPGMA cluster analYsis.'.'... -

Electronic Laboratory Reporting Use Case January 2011
ILLINOIS HEALTH INFORMATION EXCHANGE Electronic Laboratory Reporting Use Case Electronic Laboratory Reporting and Health Information Exchange Illinois Health Information Exchange Public Health Work Group January 2011 Electronic Laboratory Reporting Use Case January 2011 Table of Contents 1.0 Executive Summary……………………………………………………………………….3 2.0 Introduction…………………………………………………………………...……………..5 3.0 Scope……………………………………..………………………………………………………5 4.0 Use Case Stakeholders…………………………………………………………….….....6 5.0 Issues and Obstacles……………………………………………………………………...8 6.0 Use Case Pre-Conditions .………………….…………………………………………...8 7.0 Use Case Post-Conditions.……………………………………………………………...9 8.0 Detailed Scenarios/Technical Specifications.………………………………10 9.0 Validation and Certification………………………………………………………...12 Appendix ………………………………………………………………………………………….....13 Page 2 Electronic Laboratory Reporting Use Case January 2011 1.0 Executive Summary This Use Case is a product of the Public Health Work Group (PHWG) of the Illinois Health Information Exchange (HIE) Advisory Committee. The Illinois HIE Advisory Committee was constituted as the diverse public healthcare stakeholder body providing input and recommendations on the creation of the Illinois Health Information Exchange Authority (“the Authority”) as the Illinois vehicle for designing and implementing electronic health information exchange in Illinois. The establishment of the Authority marks the formal transition of the work of the HIE Advisory Committee and the Work Groups into alignment with the provisions of Illinois -

WO 2016/188962 Al 1 December 2016 (01.12.2016) P O P C T
(12) INTERNATIONAL APPLICATION PUBLISHED UNDER THE PATENT COOPERATION TREATY (PCT) (19) World Intellectual Property Organization International Bureau (10) International Publication Number (43) International Publication Date WO 2016/188962 Al 1 December 2016 (01.12.2016) P O P C T (51) International Patent Classification: (74) Agents: GOODFELLOW, Hugh Robin et al; Carpmaels C12Q 1/68 (2006.01) & Ransford LLP, One Southampton Row, London WC1B 5HA (GB). (21) International Application Number: PCT/EP2016/061599 (81) Designated States (unless otherwise indicated, for every kind of national protection available): AE, AG, AL, AM, (22) Date: International Filing AO, AT, AU, AZ, BA, BB, BG, BH, BN, BR, BW, BY, 23 May 20 16 (23.05.2016) BZ, CA, CH, CL, CN, CO, CR, CU, CZ, DE, DK, DM, (25) Filing Language: English DO, DZ, EC, EE, EG, ES, FI, GB, GD, GE, GH, GM, GT, HN, HR, HU, ID, IL, IN, IR, IS, JP, KE, KG, KN, KP, KR, (26) Publication Language: English KZ, LA, LC, LK, LR, LS, LU, LY, MA, MD, ME, MG, (30) Priority Data: MK, MN, MW, MX, MY, MZ, NA, NG, NI, NO, NZ, OM, 1508860.2 22 May 2015 (22.05.2015) GB PA, PE, PG, PH, PL, PT, QA, RO, RS, RU, RW, SA, SC, SD, SE, SG, SK, SL, SM, ST, SV, SY, TH, TJ, TM, TN, (71) Applicant: NATIONAL UNIVERSITY OF IRELAND, TR, TT, TZ, UA, UG, US, UZ, VC, VN, ZA, ZM, ZW. GALWAY [IE/IE]; University Road, Galway (IE). (84) Designated States (unless otherwise indicated, for every (72) Inventors: REDDINGTON, Kate Mary; Deerpack East, kind of regional protection available): ARIPO (BW, GH, Newport Road, Westport, Co. -
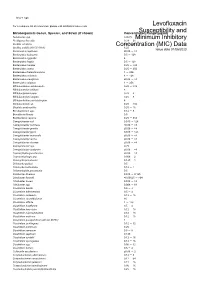
Susceptibility and Resistance Data
toku-e logo For a complete list of references, please visit antibiotics.toku-e.com Levofloxacin Microorganism Genus, Species, and Strain (if shown) Concentration Range (μg/ml)Susceptibility and Aeromonas spp. 0.0625 Minimum Inhibitory Alcaligenes faecalis 0.39 - 25 Bacillus circulans Concentration0.25 - 8 (MIC) Data Bacillus subtilis (ATCC 6051) 6.25 Issue date 01/06/2020 Bacteroides capillosus ≤0.06 - >8 Bacteroides distasonis 0.5 - 128 Bacteroides eggerthii 4 Bacteroides fragilis 0.5 - 128 Bacteroides merdae 0.25 - >32 Bacteroides ovatus 0.25 - 256 Bacteroides thetaiotaomicron 1 - 256 Bacteroides uniformis 4 - 128 Bacteroides ureolyticus ≤0.06 - >8 Bacteroides vulgatus 1 - 256 Bifidobacterium adolescentis 0.25 - >32 Bifidobacterium bifidum 8 Bifidobacterium breve 0.25 - 8 Bifidobacterium longum 0.25 - 8 Bifidobacterium pseudolongum 8 Bifidobacterium sp. 0.25 - >32 Bilophila wadsworthia 0.25 - 16 Brevibacterium spp. 0.12 - 8 Brucella melitensis 0.5 Burkholderia cepacia 0.25 - 512 Campylobacter coli 0.015 - 128 Campylobacter concisus ≤0.06 - >8 Campylobacter gracilis ≤0.06 - >8 Campylobacter jejuni 0.015 - 128 Campylobacter mucosalis ≤0.06 - >8 Campylobacter rectus ≤0.06 - >8 Campylobacter showae ≤0.06 - >8 Campylobacter spp. 0.25 Campylobacter sputorum ≤0.06 - >8 Capnocytophaga ochracea ≤0.06 - >8 Capnocytophaga spp. 0.006 - 2 Chlamydia pneumonia 0.125 - 1 Chlamydia psittaci 0.5 Chlamydia trachomatis 0.12 - 1 Chlamydophila pneumonia 0.5 Citrobacter diversus 0.015 - 0.125 Citrobacter freundii ≤0.00625 - >64 Citrobacter koseri 0.015 - -

Aquascreen® Legionella Species Qpcr Detection Kit
AquaScreen® Legionella species qPCR Detection Kit INSTRUCTIONS FOR USE FOR USE IN RESEARCH AND QUALITY CONTROL Symbols Lot No. Cat. No. Expiry date Storage temperature Number of reactions Manufacturer INDICATION The AquaScreen® Legionella species qPCR Detection kit is specifically designed for the quantitative detection of several Legionella species in water samples prepared with the AquaScreen® FastExt- ract kit. Its design complies with the requirements of AFNOR T90-471 and ISO/TS 12869:2012. Legionella are ubiquitous bacteria in surface water and moist soil, where they parasitize protozoa. The optimal growth temperature lies between +15 and +45 °C, whereas these gram-negative bacteria are dormant below 20 °C and do not survive above 60 °C. Importantly, Legionella are well-known as opportunistic intracellular human pathogens causing Legionnaires’ disease and Pontiac fever. The transmission occurs through inhalation of contami- nated aerosols generated by an infected source (e.g. human-made water systems like shower- heads, sink faucets, heaters, cooling towers, and many more). In order to efficiently prevent Legionella outbreaks, water safety control measures need syste- matic application but also reliable validation by fast Legionella testing. TEST PRINCIPLE The AquaScreen® Legionella species Kit uses qPCR for quantitative detection of legionella in wa- ter samples. In contrast to more time-consuming culture-based methods, AquaScreen® assays need less than six hours including sample preparation and qPCR to reliably detect Legionella. Moreover, the AquaScreen® qPCR assay has proven excellent performance in terms of specificity and sensitivity: other bacterial genera remain undetected whereas linear quantification is obtai- ned up to 1 x 106 particles per sample, therefore requiring no material dilution. -

LEGIONELLA in the ENVIRONMENT (The Cause of Legionnaires’ Disease)
Review of Current Knowledge ___________________________________________________________ LEGIONELLA in the ENVIRONMENT (the cause of Legionnaires’ disease) A Review of Current Knowledge FR/R0004 September 2001 © Foundation for Water Research Price: £15.00 (20% discount to FWR Members) Foundation for Water Research TECHNICAL ENQUIRIES Allen House, The Listons, Liston Road, Marlow, Any enquiries relating to this report should be Bucks SL7 1FD, U.K. addressed to:- Tele: +44(0)1628 891589 Professor D H Newsome, CBE Fax: +44(0)1628 472711 Foundation for Water Research E-mail: [email protected] _________________________________________________________________ Review of Current Knowledge ___________________________________________________________ FWR Reviews of Current Knowledge (ROCKs) focus on water-environment topics which are of broad interest both within and outside the water industry and which may form the subject of debate in the public and/or political arenas. The object of a ROCK is to provide concise, independent scientific and technical information on the subject so as to facilitate a wider understanding of the issues involved and to promote informed opinion about them © Foundation for Water Research 2001 The contents of this report are the copyright of the Foundation for Water Research and all rights are reserved. No part of this publication may be reproduced, stored in a retrieval system or transmitted, in any form or by any means electronic, mechanical, recording or otherwise, without the prior written consent of the copyright holders __________________________________________________________________ Review of Current Knowledge ___________________________________________________________ LEGIONELLA IN THE ENVIRONMENT (the cause of Legionnaires’ disease) A Review of Current Knowledge Introduction Legionnaires' disease was first recognised as a result of an outbreak of acute pneumonia that occurred at the convention of the American Legion in Philadelphia, USA during 1976. -

ATCC® Food & Water Testing Reference Strains Guide
ATCC® FOOD & WatER TESTING REFERENCE STRAINS GUIDE THE ESSENTIALS OF LIFE SCIENCE RESEARCH GLOBALLY DELIVERED™ We’ve taken cryopreservation to the next level of “cool” CoolCell® LX Container The CoolCell® LX container is the definitive cryopreservation tool, providing: • Ample sample space to create working stocks • Durable, alcohol-free construction • Dependable freezing rate An easy-to-use device designed to support cell, viral, bacterial, protozoan, and fungal cultures using a slow freezing rate of -1˚C per minute in a standard -80˚C freezer. Trust the leader in cryopreservation techniques, with over 85 years of experience, and order your CoolCell® LX Container (ATCC® No. ACS-6000™) today! CoolCell® is a registered trademark of BioCision, LLC. ATCC® is a registered trademark of the American Type Culture Collection. Table of Contents Introduction ........................................................................................................................................................... 4 ATCC Bacteria by Genus ................................................................................................................................ 5 Featured: Cronobacter sakazakii ...................................................................................................................................... 6 Non-O157 STEC Strains Panel ........................................................................................................................8 Salmonella Panel .............................................................................................................................................12 -

Legionella and Non-Tuberculous Mycobacteria Using MALDI TOF MS (Matrix Assisted Laser Desorption Ionisation Time of Flight Mass Spectrometry)
View metadata, citation and similar papers at core.ac.uk brought to you by CORE provided by OTHES DIPLOMARBEIT Titel der Diplomarbeit Establishment of a reference database for Acanthamoeba, Legionella and non-tuberculous mycobacteria using MALDI TOF MS (Matrix Assisted Laser Desorption Ionisation Time of Flight Mass Spectrometry) Verfasserin Dzenita HASANACEVIC angestrebter akademischer Grad Magistra der Naturwissenschaften (Mag.rer.nat.) Wien, 2012 Studienkennzahl lt. A 442 Studienblatt: Studienrichtung lt. Studienblatt: Anthropologie Betreuerin: Ass. Prof. Univ. Doz. Mag. Dr. Julia Walochnik Contents 1 ABBREVIATIONS ..................................................................................................... 5 2 INTRODUCTION ....................................................................................................... 6 2.1 Acanthamoeba .................................................................................................... 6 2.1.1 Classification ................................................................................................ 6 2.1.1.1 Phylogeny of Acanthamoeba ................................................................. 6 2.1.1.2 Methods of classification ....................................................................... 8 2.1.2 Ecology and geographical distribution ........................................................ 11 2.1.2.1 Life cycle ............................................................................................. 11 2.1.2.2 Trophozoites ...................................................................................... -

Legionella Human Health Criteria Document
United States Office of Science and Technology EPA-822-R-99-001 Environmental Protection Office of Water November 1999 Agency Washington, DC 20460 www.epa.gov Legionella: Human Health Criteria Document ACKNOWLEDGMENTS This document was prepared for the U. S. Environmental Protection Agency, Office of Ground Water and Drinking Water (OGWDW) by the Office of Science and Technology (OST) under contract with ICF Consulting Group (Contract No. 68-C6-0029). Overall planning and management for the preparation of this document was provided by Latisha S. Parker, MS of OST. EPA acknowledges the valuable contributions of those who reviewed this document. They include Lisa Almodovar MPH, Robin Oshiro, MS and Stephen Schaub, Ph.D. of the U.S. EPA. EPA also recognizess the following external peer reviewers for their excellent review and valuable comments on the draft document: Janette E. Stout Ph.D. and Paul H. Eldelstein M.D. TABLE OF CONTENTS I. Summary ............................................................................ I-1 II. General Information and Properties .......................................................II-1 A. History .......................................................................II-1 B. Taxonomy .....................................................................II-2 C. Microbiology, Morphology, and Ecology ............................................II-6 D. Symbiosis in Microorganisms .....................................................II-7 III. Occurrence .......................................................................