Skeletal System
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Red Tail Barracuda (Acestrorhynchus Falcatus) Ecological Risk Screening Summary
Red Tail Barracuda (Acestrorhynchus falcatus) Ecological Risk Screening Summary U.S. Fish and Wildlife Service, March 2014 Revised, January 2018 and June 2018 Web Version, 6/7/2018 Photo: S. Brosse. Licensed under Creative Commons (CC BY-NC). Available: http://www.fishbase.org/photos/PicturesSummary.php?StartRow=0&ID=23498&what=species& TotRec=2 (January 2018). 1 1 Native Range, and Status in the United States Native Range From Froese and Pauly (2017): “South America: Amazon and Orinoco River basins and rivers of Guyana, Suriname and French Guiana.” Status in the United States This species has not been reported as introduced or established in the United States. This species is in trade in the United States. For example: From Pet Zone Tropical Fish (2018): “Red Tail Barracuda […] Your Price: $29.99 […] Product Description Red Tail Barracuda (Acestrorhynchus falcatus)” Pet Zone Tropical Fish is based in San Diego, California. From Arizona Aquatic Gardens (2018): “Yellow Tail Barracuda Acestrorhynchus falcatus List: $129.00 - $149.00 $68.00 – $88.00” Arizona Aquatic Gardens is based in Tucson, Arizona. Means of Introductions in the United States This species has not been reported as introduced or established in the United States. 2 Biology and Ecology Taxonomic Hierarchy and Taxonomic Standing From ITIS (2018): Kingdom Animalia Subkingdom Bilateria Infrakingdom Deuterostomia Phylum Chordata Subphylum Vertebrata Infraphylum Gnathostomata Superclass Osteichthyes Class Actinopterygii 2 Subclass Neopterygii Infraclass Teleostei Superorder Ostariophysi -

Chapter 11 the Biology and Ecology of the Oceanic Whitetip Shark, Carcharhinus Longimanus
Chapter 11 The Biology and Ecology of the Oceanic Whitetip Shark, Carcharhinus longimanus Ramón Bonfi l, Shelley Clarke and Hideki Nakano Abstract The oceanic whitetip shark (Carcharhinus longimanus) is a common circumtropical preda- tor and is taken as bycatch in many oceanic fi sheries. This summary of its life history, dis- tribution and abundance, and fi shery-related information is supplemented with unpublished data taken during Japanese tuna research operations in the Pacifi c Ocean. Oceanic whitetips are moderately slow-growing sharks that do not appear to have differential growth rates by sex, and individuals in the Atlantic and Pacifi c Oceans seem to grow at similar rates. They reach sexual maturity at approximately 170–200 cm total length (TL), or 4–7 years of age, and have a 9- to 12-month embryonic development period. Pupping and nursery areas are thought to exist in the central Pacifi c, between 0ºN and 15ºN. According to two demographic metrics, the resilience of C. longimanus to fi shery exploitation is similar to that of blue and shortfi n mako sharks. Nevertheless, reported oceanic whitetip shark catches in several major longline fi sheries represent only a small fraction of total shark catches, and studies in the Northwest Atlantic and Gulf of Mexico suggest that this species has suffered signifi cant declines in abundance. Stock assessment has been severely hampered by the lack of species-specifi c catch data in most fi sheries, but recent implementation of species-based reporting by the International Commission for the Conservation of Atlantic Tunas (ICCAT) and some of its member countries will provide better data for quantitative assessment. -

2. Bilateral Cleft Anatomy 19
BILATERAL CLEFT ANATOMY IS ATTACHED TO THE SINGLE CLEFT THE PREMAXILLA NORMALLY ROTATED OUTWARD MAXILLA ON ONE SIDE AND THIS ENTIRE COMPONENT IS THE CLEFT SIDE MAXILLA IN AN VARYING DEGREES FROM ASYMMETRICAL DIFFERENT DISTORTION DOUBLE CLEFTS PRESENT AN ENTIRELY CONFIGURA TION IN THE COMPLETE BILATERAL CLEFT THE PREMAXILLA IS UNATTACHED THREE WHICH TO EITHER MAXILLA THUS THERE ARE SEPARATE COMPONENTS IN THEIR DISTORTION THE MAXILLAE ARE MORE OR LESS SYMMETRICAL TWO WHILE THE ARE USUALLY EQUAL TO EACH OTHER IN SIZE AND POSITION FORWARD ITS IN CENTRAL PREMAXILLARY ELEMENT PROCEEDS ON OWN WITHIN ITSELF FOR DIFFERENT DEGREES BUT WITH SYMMETRY EXCEPT IJI POSSIBLE DEVIATION FRONTONASAL THE COMPLETE SEPARATION OF THE CENTRAL COMPONENT OF PROLABIUM AND PREMAXILLA FROM THE LATERAL MAXILLARY SEGMENTS THE VASCULAR ABNORMALLY INFLUENCES NOSE PHILTRUM MUSCULATURE AND OF ALL THREE ELEMENTS ITY NERVE SUPPLY GROWTH DEVELOPMENT WHERE THE CLEFT IS INCOMPLETE ON BOTH SIDES THE DEFORMITY IS LESS AND IS STILL SYMMETRICAL IN SUCH CASE THERE IS USUALLY MORE OR LESS INTACT ALVEOLUS AND LITTLE OR NO PROTRUSION OF THE PRE THE MAXILLA THE COLUMELLA IS LIKELY TO BE LONGER THAN IN COMPLETE CLEFT BUT NOT OF NORMAL LENGTH SOMETIMES SOMETIMES THE DEGREE OF CLEFT VARIES ON EACH SIDE SIDE THE INCOMPLETENESS SHOWS AS ONLY THE SLIGHTEST NOTCH ON ONE SIDE OR THERE CLEFT ON THE OPPOSITE AND HALFWAY OR THREEQUARTER ON THE CLEFT ONE SIDE AND AN INCOMPLETE ONE CAN BE COMPLETE ON OF THE EXASPERATING ASPECT OTHER WHICH CONDITION EXAGGERATES THE ROTATION OF THE IN THE AND NOSE -

Mouth Esophagus Stomach Rectum and Anus Large Intestine Small
1 Liver The liver produces bile, which aids in digestion of fats through a dissolving process known as emulsification. In this process, bile secreted into the small intestine 4 combines with large drops of liquid fat to form Healthy tiny molecular-sized spheres. Within these spheres (micelles), pancreatic enzymes can break down fat (triglycerides) into free fatty acids. Pancreas Digestion The pancreas not only regulates blood glucose 2 levels through production of insulin, but it also manufactures enzymes necessary to break complex The digestive system consists of a long tube (alimen- 5 carbohydrates down into simple sugars (sucrases), tary canal) that varies in shape and purpose as it winds proteins into individual amino acids (proteases), and its way through the body from the mouth to the anus fats into free fatty acids (lipase). These enzymes are (see diagram). The size and shape of the digestive tract secreted into the small intestine. varies in each individual (e.g., age, size, gender, and disease state). The upper part of the GI tract includes the mouth, throat (pharynx), esophagus, and stomach. The lower Gallbladder part includes the small intestine, large intestine, The gallbladder stores bile produced in the liver appendix, and rectum. While not part of the alimentary 6 and releases it into the duodenum in varying canal, the liver, pancreas, and gallbladder are all organs concentrations. that are vital to healthy digestion. 3 Small Intestine Mouth Within the small intestine, millions of tiny finger-like When food enters the mouth, chewing breaks it 4 protrusions called villi, which are covered in hair-like down and mixes it with saliva, thus beginning the first 5 protrusions called microvilli, aid in absorption of of many steps in the digestive process. -

MAF Underwatermission Synopsis Final
Conceived by Max Serio Developed by Max Serio, John Hopkins, Martin Kase, Tina Dalton Directed by Max Serio, Tina Dalton Narrated by Rachel King, Juliet Jordan, Marcello Fabrizi Underwater Mission: Cleaner Friends First episode of the series. Our heroes: Sara,Maxi and Emma the sea turtle will explore who are their Cleaner Friends. Their adventure will be supported with the valuable information of "Sea Pad" their "friend-board computer". Cleaner Friends : Cleaner shrimp,moray eel,Blue Streak Cleaner Wrasse,Moorish Idols,Humphead Wrasse,Spadefish, sea star,Mushroom Coral,Bristletooths. Underwater Mission: Predators In this episode Sara and Max will experience an interesting trip with Emma the sea tur- tle. “Sea Pad” is going to show them the most interesting underwater predators and their habbits. Predators : Mooray eel (Ribbon eel, White eyed moray eel), Sand conger eel, Barracudas, Stonefish, Anglerfish, Lionfish, Mantis Shrimp, White tip reef shark, Tiger Shark Underwater Mission: Crazy Colours Maxi and Sara are going to visit the most colourful environment they have ever seen. Emma the sea turtle will take them to an underwater trip where they find the beautiful wolrd of crazy-coloured fish. Crazy-coloured fish : Gold Belly Damsel Fish, Emperor Angelfish, Yellow Ribbon Sweetlip, Peach Fairies, Anemones, Corals, Clown Trigger fish, Butterfly fish, Leopard coral trout, Scribbled Filefish, Lionfish, Cuttlefish, Nudibranch, Parrotfish Underwater Mission: Startling Shapes There are many shapes that the sea creatures and objects have. Emma, Sara and Maxi are going to discover as much of them as they can. Those they can’t spot on the first glance will be uncovered by the trusted clever “Sea pad”. -

Forage Fish Management Plan
Oregon Forage Fish Management Plan November 19, 2016 Oregon Department of Fish and Wildlife Marine Resources Program 2040 SE Marine Science Drive Newport, OR 97365 (541) 867-4741 http://www.dfw.state.or.us/MRP/ Oregon Department of Fish & Wildlife 1 Table of Contents Executive Summary ....................................................................................................................................... 4 Introduction .................................................................................................................................................. 6 Purpose and Need ..................................................................................................................................... 6 Federal action to protect Forage Fish (2016)............................................................................................ 7 The Oregon Marine Fisheries Management Plan Framework .................................................................. 7 Relationship to Other State Policies ......................................................................................................... 7 Public Process Developing this Plan .......................................................................................................... 8 How this Document is Organized .............................................................................................................. 8 A. Resource Analysis .................................................................................................................................... -

Study Guide Medical Terminology by Thea Liza Batan About the Author
Study Guide Medical Terminology By Thea Liza Batan About the Author Thea Liza Batan earned a Master of Science in Nursing Administration in 2007 from Xavier University in Cincinnati, Ohio. She has worked as a staff nurse, nurse instructor, and level department head. She currently works as a simulation coordinator and a free- lance writer specializing in nursing and healthcare. All terms mentioned in this text that are known to be trademarks or service marks have been appropriately capitalized. Use of a term in this text shouldn’t be regarded as affecting the validity of any trademark or service mark. Copyright © 2017 by Penn Foster, Inc. All rights reserved. No part of the material protected by this copyright may be reproduced or utilized in any form or by any means, electronic or mechanical, including photocopying, recording, or by any information storage and retrieval system, without permission in writing from the copyright owner. Requests for permission to make copies of any part of the work should be mailed to Copyright Permissions, Penn Foster, 925 Oak Street, Scranton, Pennsylvania 18515. Printed in the United States of America CONTENTS INSTRUCTIONS 1 READING ASSIGNMENTS 3 LESSON 1: THE FUNDAMENTALS OF MEDICAL TERMINOLOGY 5 LESSON 2: DIAGNOSIS, INTERVENTION, AND HUMAN BODY TERMS 28 LESSON 3: MUSCULOSKELETAL, CIRCULATORY, AND RESPIRATORY SYSTEM TERMS 44 LESSON 4: DIGESTIVE, URINARY, AND REPRODUCTIVE SYSTEM TERMS 69 LESSON 5: INTEGUMENTARY, NERVOUS, AND ENDOCRINE S YSTEM TERMS 96 SELF-CHECK ANSWERS 134 © PENN FOSTER, INC. 2017 MEDICAL TERMINOLOGY PAGE III Contents INSTRUCTIONS INTRODUCTION Welcome to your course on medical terminology. You’re taking this course because you’re most likely interested in pursuing a health and science career, which entails proficiencyincommunicatingwithhealthcareprofessionalssuchasphysicians,nurses, or dentists. -

Il/I,E,Icanjluseum
il/i,e,icanJluseum PUBLISHED BY THE AMERICAN MUSEUM OF NATURAL HISTORY CENTRAL PARK WEST AT 79TH STREET, NEW YORK 24, N.Y. NUMBER 1870 FEBRUARY 26, 1958 The Role of the "Third Eye" in Reptilian Behavior BY ROBERT C. STEBBINS1 AND RICHARD M. EAKIN2 INTRODUCTION The pineal gland remains an organ of uncertain function despite extensive research (see summaries of literature: Pflugfelder, 1957; Kitay and Altschule, 1954; and Engel and Bergmann, 1952). Its study by means of pinealectomy has been hampered in the higher vertebrates by its recessed location and association with large blood vessels which have made difficult its removal without brain injury or serious hemorrhage. Lack of purified, standardized extracts, improper or inadequate extrac- tion techniques (Quay, 1956b), and lack of suitable assay methods to test biological activity have hindered the physiological approach. It seems probable that the activity of the gland varies among different species (Engel and Bergmann, 1952), between individuals of the same species, and within the same individual. This may also have contrib- uted to the variable results obtained with pinealectomy, injection, and implantation experiments. The morphology of the pineal apparatus is discussed in detail by Tilney and Warren (1919) and Gladstone and Wakely (1940). Only a brief survey is presented here for orientation. In living vertebrates the pineal system in its most complete form may be regarded as consisting of a series of outgrowths situated above the third ventricle in the roof of the diencephalon. In sequence these outgrowths are the paraphysis, dorsal sac, parapineal, and pineal bodies. The paraphysis, the most I University of California Museum of Vertebrate Zoology. -

Comparison of Greater Palatine Nerve Block with Intravenous Fentanyl for Postoperative Analgesia Following Palatoplasty in Children
Jemds.com Original Research Article Comparison of Greater Palatine Nerve Block with Intravenous Fentanyl for Postoperative Analgesia Following Palatoplasty in Children Amol Singam1, Saranya Rallabhandi2, Tapan Dhumey3 1Department of Anaesthesiology, JNMC, DMIMS, Sawangi Meghe, Wardha Maharashtra, India. 2Department of Anaesthesiology, JNMC, DMIMS, Sawangi Meghe, Wardha, Maharashtra, India. 3Department of Anaesthesiology, JNMC, DMIMS, Sawangi Meghe, Wardha, Maharashtra, India. ABSTRACT BACKGROUND Good pain relief after palatoplasty is important as inadequate analgesia with vigorous Corresponding Author: cry leads to wound dehiscence, removal of sutures and extra nursing care. Decrease Dr. Saranya Rallabhandi, in oxygen requirement and cardio-respiratory demand occur with good pain relief Assisstant Professor, and also promotes early recovery. Preoperative opioids have concerns like sedation, Department of Anesthesiology, AVBRH, DMIMS (DU), Sawangi Meghe, respiratory depression and airway compromise. Greater palatine nerve block with Wardha- 442001, Maharashtra, India. bupivacaine is safe and effective without the risk of respiratory depression. The study E-mail: [email protected] was done to compare pain relief postoperatively with intravenous fentanyl and greater palatine nerve block in children following palatoplasty. DOI: 10.14260/jemds/2020/549 METHODS How to Cite This Article: 80 children of ASA I & II, between 1 to 7 years were included and allocated into two Singam A, Rallabhandi S, Dhumey T. Comparison of greater palatine nerve block groups of 40 each. Analgesic medication was given preoperatively after induction of with intravenous fentanyl for postoperative general anaesthesia, children in Group B received greater palatine nerve block with analgesia following palatoplasty in -1 2 mL 0.25% inj. Bupivacaine (1 mL on each side) and Group F received 2 μg Kg I.V. -

Mammals from the Mesozoic of Mongolia
Mammals from the Mesozoic of Mongolia Introduction and Simpson (1926) dcscrihed these as placental (eutherian) insectivores. 'l'he deltathcroids originally Mongolia produces one of the world's most extraordi- included with the insectivores, more recently have narily preserved assemblages of hlesozoic ma~nmals. t)een assigned to the Metatheria (Kielan-Jaworowska Unlike fossils at most Mesozoic sites, Inany of these and Nesov, 1990). For ahout 40 years these were the remains are skulls, and in some cases these are asso- only Mesozoic ~nanimalsknown from Mongolia. ciated with postcranial skeletons. Ry contrast, 'I'he next discoveries in Mongolia were made by the Mesozoic mammals at well-known sites in North Polish-Mongolian Palaeontological Expeditions America and other continents have produced less (1963-1971) initially led by Naydin Dovchin, then by complete material, usually incomplete jaws with den- Rinchen Barsbold on the Mongolian side, and Zofia titions, or isolated teeth. In addition to the rich Kielan-Jaworowska on the Polish side, Kazi~nierz samples of skulls and skeletons representing Late Koualski led the expedition in 1964. Late Cretaceous Cretaceous mam~nals,certain localities in Mongolia ma~nmalswere collected in three Gohi Desert regions: are also known for less well preserved, but important, Bayan Zag (Djadokhta Formation), Nenlegt and remains of Early Cretaceous mammals. The mammals Khulsan in the Nemegt Valley (Baruungoyot from hoth Early and Late Cretaceous intervals have Formation), and llcrmiin 'ISav, south-\vest of the increased our understanding of diversification and Neniegt Valley, in the Red beds of Hermiin 'rsav, morphologic variation in archaic mammals. which have heen regarded as a stratigraphic ecluivalent Potentially this new information has hearing on the of the Baruungoyot Formation (Gradzinslti r't crl., phylogenetic relationships among major branches of 1977). -

A Multifunction Trade-Off Has Contrasting Effects on the Evolution of Form and Function ∗ KATHERINE A
Syst. Biol. 0():1–13, 2020 © The Author(s) 2020. Published by Oxford University Press, on behalf of the Society of Systematic Biologists. All rights reserved. For permissions, please email: [email protected] DOI:10.1093/sysbio/syaa091 Downloaded from https://academic.oup.com/sysbio/advance-article/doi/10.1093/sysbio/syaa091/6040745 by University of California, Davis user on 08 January 2021 A Multifunction Trade-Off has Contrasting Effects on the Evolution of Form and Function ∗ KATHERINE A. CORN ,CHRISTOPHER M. MARTINEZ,EDWARD D. BURRESS, AND PETER C. WAINWRIGHT Department of Evolution & Ecology, University of California, Davis, 2320 Storer Hall, 1 Shields Ave, Davis, CA, 95616 USA ∗ Correspondence to be sent to: University of California, Davis, 2320 Storer Hall, 1 Shields Ave, Davis, CA 95618, USA; E-mail: [email protected] Received 27 August 2020; reviews returned 14 November 2020; accepted 19 November 2020 Associate Editor: Benoit Dayrat Abstract.—Trade-offs caused by the use of an anatomical apparatus for more than one function are thought to be an important constraint on evolution. However, whether multifunctionality suppresses diversification of biomechanical systems is challenged by recent literature showing that traits more closely tied to trade-offs evolve more rapidly. We contrast the evolutionary dynamics of feeding mechanics and morphology between fishes that exclusively capture prey with suction and multifunctional species that augment this mechanism with biting behaviors to remove attached benthic prey. Diversification of feeding kinematic traits was, on average, over 13.5 times faster in suction feeders, consistent with constraint on biters due to mechanical trade-offs between biting and suction performance. -
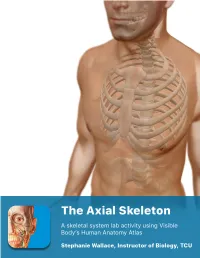
Lab Manual Axial Skeleton Atla
1 PRE-LAB EXERCISES When studying the skeletal system, the bones are often sorted into two broad categories: the axial skeleton and the appendicular skeleton. This lab focuses on the axial skeleton, which consists of the bones that form the axis of the body. The axial skeleton includes bones in the skull, vertebrae, and thoracic cage, as well as the auditory ossicles and hyoid bone. In addition to learning about all the bones of the axial skeleton, it is also important to identify some significant bone markings. Bone markings can have many shapes, including holes, round or sharp projections, and shallow or deep valleys, among others. These markings on the bones serve many purposes, including forming attachments to other bones or muscles and allowing passage of a blood vessel or nerve. It is helpful to understand the meanings of some of the more common bone marking terms. Before we get started, look up the definitions of these common bone marking terms: Canal: Condyle: Facet: Fissure: Foramen: (see Module 10.18 Foramina of Skull) Fossa: Margin: Process: Throughout this exercise, you will notice bold terms. This is meant to focus your attention on these important words. Make sure you pay attention to any bold words and know how to explain their definitions and/or where they are located. Use the following modules to guide your exploration of the axial skeleton. As you explore these bones in Visible Body’s app, also locate the bones and bone markings on any available charts, models, or specimens. You may also find it helpful to palpate bones on yourself or make drawings of the bones with the bone markings labeled.