Sequential Combination of Decitabine
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Gy Total Body Irradiation Followed by Allogeneic Hematopoietic St
Bone Marrow Transplantation (2009) 44, 785–792 & 2009 Macmillan Publishers Limited All rights reserved 0268-3369/09 $32.00 www.nature.com/bmt ORIGINAL ARTICLE Fludarabine, amsacrine, high-dose cytarabine and 12 Gy total body irradiation followed by allogeneic hematopoietic stem cell transplantation is effective in patients with relapsed or high-risk acute lymphoblastic leukemia F Zohren, A Czibere, I Bruns, R Fenk, T Schroeder, T Gra¨f, R Haas and G Kobbe Department of Haematology, Oncology and Clinical Immunology, Heinrich Heine-University, Du¨sseldorf, Germany In this prospective study, we examined the toxicity and Introduction efficacy of an intensified conditioning regimen for treatment of patients with relapsed or high-risk acute The use of intensified conventional induction chemothera- lymphoblastic leukemia who undergo allogeneic hemato- pies in the treatment of newly diagnosed ALL in adult poietic stem cell transplantation. Fifteen patients received patients has led to initial CR rates of up to 90%. But, as fludarabine 30 mg/m2, cytarabine 2000 mg/m2, amsacrine some patients are even refractory to first-line therapy, the 100 mg/m2 on days -10, -9, -8 and -7, anti-thymocyte majority of the responding patients unfortunately suffer globulin (ATG-Fresenius) 20 mg/kg body weight on days from relapse. Therefore, the probability of long-term -6, -5 and -4 and fractionated total body irradiation disease-free survival is o30%.1–5 Risk stratification based 2 Â 2 Gy on days -3, -2 and -1 (FLAMSA-ATG-TBI) on prognostic factors allows identification of high-risk before allogeneic hematopoietic stem cell transplantation. ALL patients with poor prognosis.6–12 Myeloablative At the time of hematopoietic stem cell transplantation, 10 conditioning therapy followed by allogeneic hematopoietic patients were in complete remission (8 CR1; 2 CR2), 3 stem cell transplantation (HSCT) is currently the treatment with primary refractory and 2 suffered from refractory of choice for these high-risk patients.13–15 A combination of relapse. -

CK0403, a 9‑Aminoacridine, Is a Potent Anti‑Cancer Agent in Human Breast Cancer Cells
MOLECULAR MEDICINE REPORTS 13: 933-938, 2016 CK0403, a 9‑aminoacridine, is a potent anti‑cancer agent in human breast cancer cells YUAN-WAN SUN1, KUEN-YUAN CHEN2, CHUL-HOON KWON3 and KUN-MING CHEN1 1Department of Biochemistry and Molecular Biology, College of Medicine, Pennsylvania State University, Hershey, PA 17033, USA; 2Department of Surgery, National Taiwan University Hospital, Taipei 8862, Taiwan, R.O.C.; 3Department of Pharmaceutical Sciences, College of Pharmacy and Allied Health Professions, St. John's University, New York, NY 11439, USA Received November 11, 2014; Accepted October 22, 2015 DOI: 10.3892/mmr.2015.4604 Abstract. 3-({4-[4-(Acridin-9-ylamino)phenylthio]phenyl} reduced growth inhibitory activity under hypoxic conditions, (3-hydroxypropyl)amino)propan-1-ol (CK0403) is a which can induce autophagy. Collectively, the present results sulfur-containing 9-anilinoacridine analogue of amsacrine and supported that CK0403 is highly potent and more effective than was found to be more potent than its analogue 2-({4-[4-(acridin- CK0402 against estrogen receptor-negative and 9-ylamino)phenylthio]phenyl}(2-hydroxyethyl)amino) HER2-overexpressing breast cancer cell lines, suggesting its ethan-1-ol (CK0402) and amsacrine in the inhibition of the future application for chemotherapy in breast cancer. topoisomerase II-catalyzed decatenation reaction. A previous study by our group reported that CK0402 was effective against Introduction numerous breast cancer cell lines, and the combination of CK0402 with herceptin enhanced its activity in HER2(+) Breast cancer is the most common type of cancer diagnosed SKBR-3 cells. In order to identify novel chemotherapeutic agents in American women and is the second most common cause with enhanced potency, the present study explored the potential of cancer-associated mortality. -

(Idarubicin) with Pharmacokinetic and in Vitro Drug Sensitivity Testing in Children with Refractory Leukemia1
(CANCER RESEARCH 48, 5348-5352. September 15. 1988] Phase I Clinical Trial of Orally Administered 4-Demethoxydaunorubicin (Idarubicin) with Pharmacokinetic and in Vitro Drug Sensitivity Testing in Children with Refractory Leukemia1 Ching-Hon Pui,2 Siebold S. N. de Graaf,3 Lois W. Dow, John H. Rodman, William E. Evans, Bruce S. Alpert, and Sharon B. Murphy Departments ofHematology and Oncology [C-H. P., L. W. D., S. B. M.] and Pharmaceutical Division [S. S. N. d. G., J. H. R., W. E. E.], St. Jude Children's Research Hospital, and Divisions of Hematology-Oncology [C-H. P., L. W. D., S. B. M.] and Cardiology [B. S. A.], Department of Pediatrics, University of Tennessee, Memphis, College of Medicine, Memphis, Tennessee 38101 ABSTRACT than that of analogues or i.v. idarubicin (9, 16). However, nausea and vomiting were generally more severe in patients Fifteen children with acute leukemia in relapse, refractory to conven receiving oral idarubicin (16). As a single agent in adults, oral tional therapy, were treated with idarubicin administered orally for 3 consecutive days in dosages ranging from 30 to 50 mg/m2 per day at 19- idarubicin has antileukemic activity at dosages of 45 to 90 mg/ m2 distributed over 3 consecutive days (9, 17, 18). Very little to 21-day intervals. Gastrointestinal complications, including nausea, vomiting, abdominal pain, diarrhea and stomatitis, were the major forms information is available for oral idarubicin treatment in children of dose-limiting toxicity, affecting the majority of patients at all levels of with leukemia. We report here the results of Phase I clinical, idarubicin dosage. -
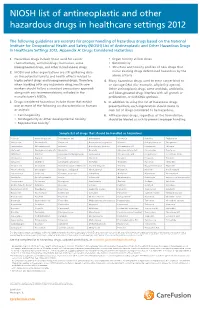
NIOSH List of Antineoplastic and Other Hazardous Drugs in Healthcare Settings 2012
NIOSH list of antineoplastic and other hazardous drugs in healthcare settings 2012 The following guidelines are excerpts for proper handling of hazardous drugs based on the National Institute for Occupational Health and Safety (NIOSH) List of Antineoplastic and Other Hazardous Drugs in Healthcare Settings 2012, Appendix A: Drugs Considered Hazardous 1. Hazardous drugs include those used for cancer • Organ toxicity at low doses* chemotherapy, antiviral drugs, hormones, some • Genotoxicity** bioengineered drugs, and other miscellaneous drugs. • Structure and toxicity profiles of new drugs that 2. NIOSH and other organizations are still gathering data mimic existing drugs determined hazardous by the on the potential toxicity and health effects related to above criteria highly potent drugs and bioengineered drugs. Therefore, 4. Many hazardous drugs used to treat cancer bind to when working with any hazardous drug, health care or damage DNA (for example, alkylating agents). workers should follow a standard precautions approach Other antineoplastic drugs, some antivirals, antibiotics, along with any recommendations included in the and bioengineered drugs interfere with cell growth or manufacturer’s MSDSs. proliferation, or with DNA synthesis. 3. Drugs considered hazardous include those that exhibit 5. In addition to using the list of hazardous drugs one or more of the following six characteristics in humans presented here, each organization should create its or animals: own list of drugs considered to be hazardous. • Carcinogenicity 6. All hazardous -

Intercalating TOP2 Poisons Attenuate Topoisomerase Action at Higher Concentrations
Molecular Pharmacology Fast Forward. Published on August 9, 2019 as DOI: 10.1124/mol.119.117259 This article has not been copyedited and formatted. The final version may differ from this version. Intercalating TOP2 poisons attenuate topoisomerase action at higher concentrations Mandeep Atwal, Rebecca L. Swan, Chloe Rowe, Ka C. Lee, David C Lee, Lyle Armstrong, Ian G Cowell*, Caroline A Austin* Running title: Inhibition by doxorubicin, epirubicin and mitoxantrone MA, RLS, CR, KCL, IGC, CAA Institute for Cell and Molecular Biosciences, Newcastle University, Newcastle upon Tyne. NE2 4HH. U.K. Downloaded from DCL, LA Institute of Genetic Medicine, Newcastle University, International Centre for Life, Central Parkway, Newcastle upon Tyne. NE1 3BZ. DCL: https://orcid.org/0000-0001-6857-624X IGC: https://orcid.org/0000-0002-8606-0447 molpharm.aspetjournals.org CAA: https://orcid.org/0000-0002-1921-5947 *Corresponding authors: Ian G Cowell & Caroline A Austin Institute for Cell and Molecular Biosciences, Newcastle University, Newcastle upon Tyne NE2 4HH. United Kingdom at ASPET Journals on September 26, 2021 Tel: +44 (0)191 208 7677 Fax: +44 (0)191 208 7424 Email: [email protected], [email protected] 30 text pages (including refs and figure legends) 8 figures 0 tables 60 number of references 219 words in abstract 641 words in Introduction 1268 words in Discussion Abbreviations DMSO Dimethyl sulfoxide DOX Doxorubicin DSB Double-strand break EPI Epirubicin ICRF-193 4-[2-(3,5-Dioxo-1-piperazinyl)-1-methylpropyl]piperazine-2,6-dione TARDIS Trapped in Agarose DNA Immunostaining assay TOP2 Topoisomerase II TOP2A Topoisomerase IIα TOP2B Topoisomerase IIβ TOP2-CC Topoisomerase II covalent complex 1 Molecular Pharmacology Fast Forward. -

NIOSH List of Antineoplastic and Other Hazardous Drugs in Healthcare Settings 2010
NIOSH List of Antineoplastic and Other Hazardous Drugs in Healthcare Settings 2010 DEPARTMENT OF HEALTH AND HUMAN SERVICES Centers for Disease Control and Prevention National Institute for Occupational Safety and Health NIOSH List of Antineoplastic and Other Hazardous Drugs in Healthcare Settings 2010 DEPARTMENT OF HEALTH AND HUMAN SERVICES Centers for Disease Control and Prevention National Institute for Occupational Safety and Health This document is in the public domain and may be freely copied or reprinted. DISCLAIMER Mention of any company or product does not constitute endorsement by the National Institute for Occupational Safety and Health (NIOSH). In addition, citations to Web sites external to NIOSH do not constitute NIOSH endorsement of the sponsoring organizations or their programs or products. Furthermore, NIOSH is not responsible for the content of these Web sites. ORDERING INFORMATION To receive documents or other information about occupational safety and health topics, contact NIOSH at Telephone: 1–800–CDC–INFO (1–800–232–4636) TTY:1–888–232–6348 E-mail: [email protected] or visit the NIOSH Web site at www.cdc.gov/niosh For a monthly update on news at NIOSH, subscribe to NIOSH eNews by visiting www.cdc.gov/niosh/eNews. DHHS (NIOSH) Publication Number 2010−167 September 2010 Preamble: The National Institute for Occupational Safety and Health (NIOSH) Alert: Preventing Occupational Exposures to Antineoplastic and Other Hazardous Drugs in Health Care Settings was published in September 2004 (http://www.cdc.gov/niosh/docs/2004-165/). In Appendix A of the Alert, NIOSH identified a sample list of major hazardous drugs. The list was compiled from infor- mation provided by four institutions that have generated lists of hazardous drugs for their respec- tive facilities and by the Pharmaceutical Research and Manufacturers of America (PhRMA) from the American Hospital Formulary Service Drug Information (AHFS DI) monographs [ASHP/ AHFS DI 2003]. -

Evidence Tables Cardiomyopathy Surveillance
Who needs surveillance? 1. What is the evidence behind the conversion score for different derivates for anthracyclines (including mitoxantrone) First Author Study Design Participants Treatment Main outcomes Addt’l remarks Year Treatment era Years of follow-up van der Pal1 Retrospective cohort 5-yr survivors (N=1362) Conversion score: Refs: 2012 Doxorubicin : 1.0 Mertens (2008): late mortality 1966-1996 Daunorubicin: 1.0 Le Deley (2003): SMN after solid Epirubicin: 0.67 CA 22.2 yrs (5.0-44.5) Perez (1991): Breast CA (epi vs.dox) Mulrooney2 Retrospective cohort 5-yr Survivors (N=14, Conversion score: Conversion score based on a 2009 1970-1986 358) Doxorubicin = Daunorubicin review paper recommendations 27.0 yrs (8-51) Idarubicin = 3x doxorubicin (Pai Nahata 2000) Siblings (N=3899) Blanco3 Case-Control Case (CHF) – N=170 Conversion score: COG LTFU Conversion score based on: 2012 Control (none) – N=317 Guidelines Lehmann (2000), which is based 1966-2008 Doxorubicin: 1.0 Doxorubicin: 1.0 on sited review literature with 1 Daunorubicin: 0.75 Daunorubicin: in vivo model of acute toxicity Cases: 9.2 (0.1-35.1) 0.83 Controls: 12.3 (0.4- Epirubicin: 0.75 Epirubicin: 0.67 40) Idarubicin: 3 Idarubicin: 5 Mitoxantrone: 3 Mitoxantrone: 4 Temming4 Retrospective cohort 124/158 available for AML 10 and 12 trials Anthracycline dose range similar 2011 N=124, 86 Cardiotox analysis across AML 10 and 12, unable 86 data for late Anthracyclines: to assess dose-association 1987-2004 cardiotox Dauno and Mitox (1:5 conversion) No discussion on conversion 7.3 yrs (0-21.7) 550-610 mg/m2 factor Creutzig5 Retrospective cohort Eligible: N=1207 AML BFM 93 98 2007 1993-2003 Late Cartox eval: BFM98: 3.6ys (0.8- N=547 (45%) Dauno : Ida 1:5 7.0) 76% of echo w/in first Dauno : Mitox 1:5 BFM93: 7.5ys (1.1- 5yrs 11) van Dalen6 Systematic review Different anthracycline Dox Epi vs. -

Bc Cancer Chemotherapy Preparation and Stability
BC CANCER CHEMOTHERAPY PREPARATION AND STABILITY CHART DRUG & STRENGTH Reconstitute To Give: Vial Product Product Stability Special (Storage Prior to Use, With: Stability (for IV bag size selection, Precautions/Notes Manufacturer, Preservative see Notes†) Status) AGS-16C3F 30 mg 5.1 mL SWI1 6 mg/mL1 discard unused ≥ 0.3 mg/mL1 complete - unopened vials (Astellas) portion1 administration within may be kept at RT (F)(PFL) swirl gently; do NOT 100 mL D5W2,3 6 h RT of for up to 4h prior to do not shake shake1 **(PFL)1 reconstitution1 use if protected no preservative1 mix by gentle from light1 allow foam to clear inversion1 **(PFL) before proceeding1 record time of reconstitution Aldesleukin 22 million units 1.2 mL SWI4 18 million unit/mL 48 h F, RT4 30-70 mcg/mL4 48 h F, RT4 - do NOT use in- (1.3 mg) (1.1 mg/mL)4 line filter4 (SteriMax) direct diluent against 50 mL D5W4 - avoid (F)(PFL) side of vial during bacteriostatic water no preservative4 reconstitution4 <30 mcg/mL: for injection or NS dilute in D5W due to increased do NOT shake4 containing human aggregation4 albumin 0.1%5 - bring to room temperature prior to infusion4 SC syringe6,7 14 d F7 **(PFL) BC Cancer Chemotherapy Preparation and Stability Chart© version 2.00. All rights reserved. 1/60 This document may not be reproduced in any form without the express written permission of BC Cancer Provincial Pharmacy. Activation Date: 2 March 2006 Revised Date: 1 September 2021 BC CANCER CHEMOTHERAPY PREPARATION AND STABILITY CHART DRUG & STRENGTH Reconstitute To Give: Vial Product Product -

Chemotherapy Drugs
CHEMOTHERAPY, DRUGS, AGENTS- This document contains three lists: 1) Individual drugs/agents 2) Known protocols 3) Old protocol descriptions Each table has the name of the drug, the code by which it is represented in the database, and the name by which this drug appears in the database, if the drug can be known by more than one name. 1. Individual drugs/agents Drug Name (synonym) Code Name in database 2-CdA 63 2-CdA 4-demethoxydaunorubicin 18 Idarubicine 5- Fluorouracil 20 Fluorouracil 5FU 20 Fluorouracil 6-mercaptopurine 22 Mercaptapur6 6-MP 22 Mercaptapur6 Acridinyl anisidide 2 Amsacrine Actemra 679 Tocilizumab/ Actemra Actinomycin D 65 Dactinomycin Acyclovir 311 Acyclovir Adcetris 666 Brentuximab (Adcetris) Adriamycine 1 Adriamycine Adriblastin 1 Adriamycine Advagraf 52 Tacrolimus Agrylin 351 Anagrelide Aldesleukin 206 IL-2 Alemtuzumab 632 Campath (CD52, Alemtuzumab) Alexan 3 ARA-C / Cytarabine ALG 35 ALG Alkeran 21 Melphalan AMD-3100 36 Plerixafor Amethopterin 23 Methotrexate Amikin 326 Amikin AML like therapy 43 AML like therapy AMSA P-D 2 Amsacrine Amsacrine 2 Amsacrine Anagrelide 351 Anagrelide Analgesics 88 Analgesics Androgens 39 Androgens Anthracycline 57 Anthracycline Anti IL6 643 Anti IL6 anti rIL2 (CD25) 636 anti rIL2 (CD25) Anti tumour necrosis factor (TNF) 639 Anti tumour necrosis factor (TNF) Anti-GD2 644 Anti-GD2 antiLFA1 635 antiLFA1 Antilymphocyte globulin 35 ALG Anti-malarial 56 Anti-malarial Antimetabolites 93 Antimetabolites Antithymocyte globulin 34 ATG ARA-C 3 ARA-C Aranesp (Darbepoetin) 85 Aranesp (Darbepoetin) -

Pemetrexed and Gemcitabine As Combination Therapy for the Treatment of Group3 Medulloblastoma
Cancer Cell Article Pemetrexed and Gemcitabine as Combination Therapy for the Treatment of Group3 Medulloblastoma Marie Morfouace,1 Anang Shelat,2 Megan Jacus,3 Burgess B. Freeman III,4 David Turner,3 Sarah Robinson,1 Frederique Zindy,1 Yong-Dong Wang,5 David Finkelstein,5 Olivier Ayrault,6 Laure Bihannic,6 Stephanie Puget,7 Xiao-Nan Li,8 James M. Olson,9 Giles W. Robinson,10 R. Kiplin Guy,2 Clinton F. Stewart,3 Amar Gajjar,10 and Martine F. Roussel1,* 1Department of Tumor Cell Biology 2Department of Chemical Biology and Therapeutics 3Department of Pharmaceutical Sciences 4Department of Preclinical Pharmacokinetics 5Department of Computational Biology St. Jude Children’s Research Hospital, Memphis, TN 38105, USA 6Institut Curie/CNRS UMR 3306/INSERM U1005 Building 110, Centre Universitaire, 91405 Orsay, Cedex, France 7AP-HP, Department of Neurosurgery, Necker-Enfants Malades Hospital, Universite´ Rene Descartes, 75015 Paris, France 8Department of Pediatrics, Section of Hematology-Oncology, Baylor College of Medicine, Houston, TX 77030, USA 9Department of Pediatric Hematology-Oncology, Fred Hutchinson Cancer Research Center, Seattle, WA 98109, USA 10Department of NeuroOncology, St. Jude Children’s Research Hospital, Memphis, TN 38105, USA *Correspondence: [email protected] http://dx.doi.org/10.1016/j.ccr.2014.02.009 SUMMARY We devised a high-throughput, cell-based assay to identify compounds to treat Group3 medulloblastoma (G3 MB). Mouse G3 MBs neurospheres were screened against a library of approximately 7,000 compounds including US Food and Drug Administration-approved drugs. We found that pemetrexed and gemcitabine preferentially inhibited G3 MB proliferation in vitro compared to control neurospheres and substantially in- hibited G3 MB proliferation in vivo. -

Liste Des Molécules
Molécules Amobarbital sodium Etomidate Ketamine hydrochloride Methohexital sodium Pentobarbital sodium Pentobarbital sodium Propofol Thiopental sodium Bupivacaine hydrochloride Cocaine hydrochloride Levobupivacaine hydrochloride Lidocaine hydrochloride Lidocaine hydrochloride Lidocaine hydrochloride Procaine hydrochloride Ropivacain hydrochloride Tetracain hydrochloride Tetracain hydrochloride Tetracaine Acetylsalicylic Acid Acetylsalicylic acid lysinate Alfentanil hydrochloride Buprenorphine Buprenorphine hydrochloride Butorphanol tartrate Codeine phosphate Dezocine Diamorphine hydrochloride Fentanyl citrate Hydromorphone hydrochloride Meptazinol Metamizol sodium Methadone hydrochloride Methadone hydrochloride Morphine hydrochloride Morphine sulfate Morphine sulfate Morphine sulfate Morphine tartrate Nefopam 1/{nb} Oxycodone hydrochloride Oxycodone hydrochloride Paracetamol Paracetamol Paracetamol Parecoxib sodium Pethidine hydrochloride Piritramide Remifentanil hydrochloride Sufentanil citrate Tramadol hydrochloride Tramadol hydrochloride Ziconotide acetate Memantine Allopurinol Allopurinol sodium Tranexamic acid Tranexamic acid Amiodarone hydrochloride Amiodarone hydrochloride Amiodarone hydrochloride Diltiazem hydrochloride Disopyramide phosphate Flecainide acetate Flecainide acetate Metoprolol tartrate Metoprolol tartrate Mexiletine Nadolol Procainamide hydrochloride Procainamide hydrochloride Propafenone hydrochloride Propafenone Hydrochloride Quinidine gluconate Quinidine sulfate Verapamil hydrochloride Amikacin sulfate Amoxicillin -

Chemotherapy Drugs
CHEMOTHERAPY, DRUGS, AGENTS- This document contains three lists: 1) Individual drugs/agents 2) Known protocols 3) Old protocol descriptions Each table has the name of the drug, the code by which it is represented in the database, and the name by which this drug appears in the database if the drug can be known by more than one name. 1. Individual drugs/agents Drug Name (synonym) Code Name in database 2-CdA 63 2-CdA 4-demethoxydaunorubicin 18 Idarubicine 5- Fluorouracil 20 Fluorouracil 5FU 20 Fluorouracil 6-mercaptopurine 22 Mercaptapur6 6-MP 22 Mercaptapur6 Acridinyl anisidide 2 Amsacrine Actinomycin D 65 Dactinomycin Acyclovir 311 Acyclovir Adcetris 666 Brentuximab (Adcetris) Adriamycine 1 Adriamycine Adriblastin 1 Adriamycine Advagraf 52 Tacrolimus Agrylin 351 Anagrelide Aldesleukin 206 IL-2 Alentuzumab 632 Campath (CD52, Alentuzumab) Alexan 3 ARA-C / Cytarabine ALG 35 ALG Alkeran 21 Melphalan AMD-3100 36 Plerixafor Amethopterin 23 Methotrexate Amikin 326 Amikin AML like therapy 43 AML like therapy AMSA P-D 2 Amsacrine Amsacrine 2 Amsacrine Anagrelide 351 Anagrelide Analgesics 88 Analgesics Androgens 39 Androgens Anthracycline 57 Anthracycline Anti IL6 643 Anti IL6 anti rIL2 (CD25) 636 anti rIL2 (CD25) Anti tumour necrosis factor (TNF) 639 Anti tumour necrosis factor (TNF) Anti-GD2 644 Anti-GD2 antiLFA1 635 antiLFA1 Antilymphocyte globulin 35 ALG Anti-malarial 56 Anti-malarial Antimetabolites 93 Antimetabolites Antithymocyte globulin 34 ATG ARA-C 3 ARA-C Aranesp (Darbepoetin) 85 Aranesp (Darbepoetin) Aredia 82 Pamidronate Arranon 342