Np14-4-Tekst.Pdf
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

XENOPHORIDAE Een Overzicht Van Alle Recent Voorkomende Soorten
XENOPHORIDAE Een overzicht van alle recent voorkomende soorten. J.Goud en G.Kronenberg TROQUE (Trochus) Troque agglutinant INLEIDING Dit artikel is bedoeld om het determineren van de videerd (Ponder, 1983), waarmee nu een duide diverse soorten behorende tot de familie Xenop li jk overzicht van de familie voorhanden is. De horidae Ph il ippi, 1853 te vergemakkelijken. Tot indeling die Ponder geeft zal hier grotendeels voor kort was dit een schier ondoenlijk werk, gevolgd worden. mede door de verschillende namen die aan één Tevens betreft dit artikel de inlossing van een soort gegeven werden (synoniemen) , maar ook oude belofte om een dergelijk overzicht in Vi ta vaak dezelfde naam aan verschillende soorten Marina te publiceren (Faber, 1978). In dat artikel (homoniemen), waardoor de nomenclatuur van wordt beschreven hoe de aanhechting van ma deze groep een grote chaos was. teriaal in zi jn werk gaat en wordt ingegaan op de Enkele jaren geleden is deze groep echter gere- diverse theorieën die bestaan over het waarom vita marina zeebiologische dokumentatie juli-dec. 1988 bu ikpotigen 457 van dit opvallende gedrag. Verder worden er familie: enkele opmerkingen gemaakt over de levenswij Xenophoridae Troschel, 1852 ze en de manier van voortbeweging. Hierop zal (op lijst ICNZ opninie 715 als Philippi, 1853) verder niet worden ingegaan, wij verwijzen daar geslacht: voor naar bedoeld artikel. Xenophora Fischer von Waldheim, 1807 Uitgestorven soorten zijn in onderstaand over ondergeslacht: zicht niet opgenomen. Xenophora s.S. O. soorten : conchyliophora (Born, 1780) 0.000 OORSPRONG EN VERSPRE IDING cerea (Reeve, 1845) 0.010 flindersi flindersi (eotton & Godfrey, 1938) De Xenophoridae kennen een rijke fossiele histo 0.020 e rie. -

Report on Vanuatu's Draft National Ocean Policy Consultations
Report on Vanuatu’s Draft National Ocean Policy Consultations Our Ocean Our Culture Our People 2 | Report on Vanuatu’s Draft National Ocean Policy Consultations 2016 Report on Vanuatu’s Consultations regarding the Draft National Ocean Policy As at 27 April 2016 By the Ocean Sub Committee of the National Committee on Maritime Boundary Delimitation, Ministry of Foreign Affairs Acknowledgements This document has been prepared by the Ocean Sub Committee of the National Committee for Maritime Boundary Delimitation with the assistance of the Ministry of Tourism. We thank the MACBIO project (implemented by GIZ with technical support from IUCN and SPREP; funded by BMUB) for their support. We thank the government staff who contributed to the National Consultations, Live and Learn Vanuatu for their administrative support. We are especially grateful to the communities, government staff and other stakeholders throughout the country who contributed their ideas and opinions to help ensure the future of Vanuatu’s ocean. MACBIO Marine and Coastal Biodiversity Management in Pacific Island Countries Report on Vanuatu’s Draft National Ocean Policy Consultations 2016 | 3 Table of Contents EXECUTIVE SUMMARY __________________________________________________________________________6 1 Introduction _______________________________________________________________________________ 7 2 Methods ___________________________________________________________________________________8 3 Using the input from consultations ________________________________________________________ -

Subject/ Area: Vanuatu at the Speed We Cruise, It Will Take Us More Than
Subject/ Area: Vanuatu At the speed we cruise, it will take us more than one season to cover Vanuatu! During this past 4 months, we explored the Southern part of Vanuatu: Tanna, Aniwa, Erromango and Efate. The ultimate cruising guide for Vanuatu is the Rocket Guide (nicknamed Tusker guide, from the first sponsor - www.cruising-vanuatu.com). With charts, aerial photos and sailing directions to most anchorages, you will have no problem making landings. We also used Bob Tiews & Thalia Hearns Vanuatu cruising guide and Miz Mae’s Vanuatu guide. Those 3 reference guides and previous letters in the SSCA bulletins will help you planning a great time in Vanuatu! CM 93 electronic charts are slightly off so do not rely blindly on them! At time of writing, 100 vatu (vt) was about $1 US. Tanna: Having an official port of entry, this island was our first landfall, as cruising NW to see the Northern islands will be easier than the other way around! Port Resolution: We arrived in Port Resolution early on Lucky Thursday…lucky because that is the day of the week that the Customs and Immigration officials come the 2 1/2 hour, 4-wheel drive across from Lenakel. We checked in at no extra cost, and avoided the expense of hiring a transport (2000 vatu RT). We met Werry, the caretaker of the Port Resolution “yacht club”, donated a weary Belgian flag for his collection, and found out about the volcano visit, tours, and activities. Stanley, the son of the Chief, is responsible for relations with the yachts, and he is the tour guide or coordinator of the tours that yachties decide to do. -

Cook Islands
PACIFIC PORTS DIRECTORY 2014 Port Information List Country Ports Page AMERICAN SAMOA 5 Pago Pago 8 COOK ISLANDS 11 Arutanga (Aitutaki) 14 Avatiu (Rarotonga) 17 FIJI 21 Malau (Labasa) 24 Lautoka 26 Levuka 28 Savusavu 29 Suva 32 FRENCH POLYNESIA 37 Bora-Bora 40 Papeete 41 FEDERATED STATES OF MICRONESIA 45 Chuuk 48 Kosrae 50 Pohnpei 51 Yap 53 GUAM 57 Apra 60 KIRIBATI 65 Betio (Tarawa) 68 MARSHALL ISLANDS 71 Kwajalein 74 Majuro 75 NAURU 79 Nauru 81 NEW CALEDONIA 85 Babouillat 88 Baie Ugue 89 Kouaoua 91 1 PACIFIC PORTS DIRECTORY 2014 Country Ports Page Nepoui 92 Noumea 94 Poro 96 Thio 97 PALAU 101 Malakal 104 PAPUA NEW GUINEA 107 Aitape 118 Alotau 119 Buka 120 Daru 121 Kavieng 123 Kieta 124 Kimbe 126 Lae 127 Lorengau 129 Madang 130 Oro Bay 132 Port Moresby 133 Rabaul 134 Vanimo 136 Wewak 137 SAMOA 141 Apia 143 SOLOMON ISLANDS 147 Allardyce Harbour 150 Aola Bay 151 Gizo 152 Honiara 154 Malloco Bay 155 Noro 157 Tonga 161 ss Nuku’alofa 164 2 PACIFIC PORTS DIRECTORY 2014 Country Ports Page Pangai 165 Port Neiafu (Vava;u) 167 TUVALU 171 Funafuti 173 VANUATU 177 Port Vila 180 Port Luganville (Santo) 182 WALLIS & FUTUNA 185 Leava 188 Mata’Utu 189 Information on Quarantine can be viewed on page 192 Fiji Ports CorPoration Limited invites you to Fiji We provide: • Competitive Tariff Structure • Efficient cargo handling • Safe port and anchorage • Reliable, and friendly ports service • Cost effective storage facilities • Excellent wharf infrastructure FIJI PORTS CORPORATION LIMITED Muaiwalu House, Lot 1 Tofua street, Walu Bay, Suva P O Box 780, Suva, Fiji Island • Telephone: (679) 331 2700 Facsimile: (679) 330 0064 • Email: [email protected] Web: http://www.fijiports.com.fj 3 PACIFIC PORTS DIRECTORY 2014 4 PACIFIC PORTS DIRECTORY 2014 AMERICAN SAMOA Subject Information 1 Location 14 deg 18 min South 170 deg 42 min West 2 Capital city Pago Pago (Tutuila Island) 3 Currency US dollar (US$). -

ABSTRACT Title of Dissertation: PATTERNS IN
ABSTRACT Title of Dissertation: PATTERNS IN DIVERSITY AND DISTRIBUTION OF BENTHIC MOLLUSCS ALONG A DEPTH GRADIENT IN THE BAHAMAS Michael Joseph Dowgiallo, Doctor of Philosophy, 2004 Dissertation directed by: Professor Marjorie L. Reaka-Kudla Department of Biology, UMCP Species richness and abundance of benthic bivalve and gastropod molluscs was determined over a depth gradient of 5 - 244 m at Lee Stocking Island, Bahamas by deploying replicate benthic collectors at five sites at 5 m, 14 m, 46 m, 153 m, and 244 m for six months beginning in December 1993. A total of 773 individual molluscs comprising at least 72 taxa were retrieved from the collectors. Analysis of the molluscan fauna that colonized the collectors showed overwhelmingly higher abundance and diversity at the 5 m, 14 m, and 46 m sites as compared to the deeper sites at 153 m and 244 m. Irradiance, temperature, and habitat heterogeneity all declined with depth, coincident with declines in the abundance and diversity of the molluscs. Herbivorous modes of feeding predominated (52%) and carnivorous modes of feeding were common (44%) over the range of depths studied at Lee Stocking Island, but mode of feeding did not change significantly over depth. One bivalve and one gastropod species showed a significant decline in body size with increasing depth. Analysis of data for 960 species of gastropod molluscs from the Western Atlantic Gastropod Database of the Academy of Natural Sciences (ANS) that have ranges including the Bahamas showed a positive correlation between body size of species of gastropods and their geographic ranges. There was also a positive correlation between depth range and the size of the geographic range. -

The Genital Systems of Trivia Arctica and Trivia Monacha (Prosobranchia, Mesogastropoda) and Tributyltin Induced Imposex1
Zoo1. Beitr. N. F. 34 (3): 349 - 374 (1992) (Institut für Spezielle Zoologie und Vergleichende Embryologie der Westfälischen Wilhelms-Universität Münster) The genital systems of Trivia arctica and Trivia monacha (Prosobranchia, Mesogastropoda) and tributyltin induced imposex1 EBERHARD STROBEN, CHRISTA BROlVlMEL, JORG OEHLMANN and PIO FIORONI Received June 17 , 1992 Summary The mesogastropod prosobranchs Trivia arctica and T. monacha collected along the coast of Brittany and Normandy between 1988 and 1992 exhibit imposex or pseudohermaphroditism (occurrence of male parts in addition to the female genital system) in response to tributyltin (TBT) pollution. Beside some normal females (stage 0) different imposex stages according to the classification of FIORONI et a1. (1991) (3 a, 3 band 4 for T. arctica and 3b and 4 for T. monacha) (Fig. 1) can be distinguished and are documented with scanning electron micrographs for the first time. Additional al terations of the genital tract e.g. excrescences on the vas deferens, a coiled oviduct, a bi- or trifurcated penis or several penes (2 - 5) are described. Neither TBT-induced sterilization nor sex change occur. TBT accumulation in soft parts shows sex-related differences. The VDS (vas deferens sequence) index, uncubed RPS (relative penis size) index and average female penis length of a population are dependent from TBT con centrations in ambient sea water and TBT body burden. A statistical study, based on an analysis of natural populations of T. arctica, T. monacha and Nucella lapillus allows a comparison of the specific TBT sensitivity of the three TBT bioindicators. N. lapillus exhibits a lower threshold for imposex development and a higher TBT sen sitivity, but both Trivia species proved to be suitable TBT bioindicators. -

Fossil Flora and Fauna of Bosnia and Herzegovina D Ela
FOSSIL FLORA AND FAUNA OF BOSNIA AND HERZEGOVINA D ELA Odjeljenje tehničkih nauka Knjiga 10/1 FOSILNA FLORA I FAUNA BOSNE I HERCEGOVINE Ivan Soklić DOI: 10.5644/D2019.89 MONOGRAPHS VOLUME LXXXIX Department of Technical Sciences Volume 10/1 FOSSIL FLORA AND FAUNA OF BOSNIA AND HERZEGOVINA Ivan Soklić Ivan Soklić – Fossil Flora and Fauna of Bosnia and Herzegovina Original title: Fosilna flora i fauna Bosne i Hercegovine, Sarajevo, Akademija nauka i umjetnosti Bosne i Hercegovine, 2001. Publisher Academy of Sciences and Arts of Bosnia and Herzegovina For the Publisher Academician Miloš Trifković Reviewers Dragoljub B. Đorđević Ivan Markešić Editor Enver Mandžić Translation Amra Gadžo Proofreading Amra Gadžo Correction Sabina Vejzagić DTP Zoran Buletić Print Dobra knjiga Sarajevo Circulation 200 Sarajevo 2019 CIP - Katalogizacija u publikaciji Nacionalna i univerzitetska biblioteka Bosne i Hercegovine, Sarajevo 57.07(497.6) SOKLIĆ, Ivan Fossil flora and fauna of Bosnia and Herzegovina / Ivan Soklić ; [translation Amra Gadžo]. - Sarajevo : Academy of Sciences and Arts of Bosnia and Herzegovina = Akademija nauka i umjetnosti Bosne i Hercegovine, 2019. - 861 str. : ilustr. ; 25 cm. - (Monographs / Academy of Sciences and Arts of Bosnia and Herzegovina ; vol. 89. Department of Technical Sciences ; vol. 10/1) Prijevod djela: Fosilna flora i fauna Bosne i Hercegovine. - Na spor. nasl. str.: Fosilna flora i fauna Bosne i Hercegovine. - Bibliografija: str. 711-740. - Registri. ISBN 9958-501-11-2 COBISS/BIH-ID 8839174 CONTENTS FOREWORD ........................................................................................................... -

Pleistocene Molluscs from the Namaqualand Coast
ANNALS OF THE SOUTH AFRICAN MUSEUM ANNALE VAN DIE SUID-AFRIKAANSE MUSEUM Volume 52 Band July 1969 Julie Part 9 Dee! PLEISTOCENE MOLLUSCS FROM THE NAMAQUALAND COAST By A.J.CARRINGTON & B.F.KENSLEY are issued in parts at irregular intervals as material becomes available Obtainable from the South African Museum, P.O. Box 61, Cape Town word uitgegee in dele opongereelde tye na beskikbaarheid van stof OUT OF PRINT/UIT nRUK I, 2(1, 3, 5, 7-8), 3(1-2, 5, t.-p.i.), 5(2, 5, 7-9), 6(1, t.-p.i.), 7(1, 3), 8, 9(1-2), 10(1-3), 11(1-2, 7, t.-p.i.), 21, 24(2), 27, 31(1-3), 38, 44(4)· Price of this part/Prys van hierdie deel Rg.oo Trustees of the South African Museum © 1969 Printed in South Africa by In Suid-Afrika gedruk deur The Rustica Press, Pty., Ltd. Die Rustica-pers, Edms., Bpk. Court Road, Wynberg, Cape Courtweg, Wynberg, Kaap By A. ]. CARRINGTON & B. F. KENSLEY South African Museum, Cape Town (With plates 18 to 29 and I I figures) PAGE Introduction 189 Succession 190 Systematic discussion. 191 Acknowledgements 222 Summary. 222 References 223 INTRODUCTION In the course of an examination of the Tertiary to Recent sediments of the Namaqualand coast, being carried out by one of the authors (A.].C.), a collection of fossil molluscs was assembled from the Pleistocene horizons encountered in the area. The purpose of this paper is to introduce and describe some twenty species from this collection, including forms new to the South Mrican palaeontological literature. -

The Battle of the Coral Sea
Copy N~ 71 COMBAT NARRATIVES The Battle of the Coral Sea Consisting of the actions at T ttlagi, May 4th,· off Misima, May 7th,· and in the Coral Sea on May 8thJ 1942 British Secret . ( t::.;+.:ti~·""' lRARE . ~ J ~7 3 . C63 C6 c . 3 H. '1 -~~lFT! TE \N D :. 0 ':'~1nY T~~:IS PUBLICATION FL'~_-..s;-, :H·~:Cn-~~ .1.'0 Op-16-U, R001.13lH5, , lLl VY JJUlLDI. G. ----------~------~------~~~----~ · -- - -- m, ",, )"' tffil ,.· " Fttt1"' httJ,,, f¥¥¥l,,, f¥¥¥l,,, f¥¥¥l," t¥¥ii ·~· httl~ "1 Ill 1,,, 1••• .,, ~~~;~·;·;~ ~mn ,~:·;~7 ~ Mulro SbiJna&Uo Nishino Shima.· .· Og G to 1 . ~~t\P. ::Halw.~':"::r~ttoun • Compiled from the latest information to J Ch>cl» I ·• Kita.Io Shima Natural Scale 1·2 1,912,992 at La l. Kazan Retto . o· (Vol c ano Is) ,·, ·Io Shima '··' Minami lo Shima -~·) :Mareus receVdu. '''.Asuncitm. , ,.A.gciluu:>. ,:Pagan ;'.Alamagan. MARIANAS '-' :•:. Sarigu.an .~ ·. ·· ·1.'Farallrm.de Memnilla Tinia.n l Saipan ,Rota <fGua.m. Uli.th:i.Is ~;:·. ·:;•:Fais TORRE.$ SfR ... · ·.li . c:wessel.,.B ~tJrt- ,G. .A=hem. Torresis f!"; 0 CORAL SEA VanuaL;.;,..:o·· Bllllksls Ga.u.a.P · ;:.. EspirituSantp .\'u.: .,.S .Annra. FIJI '1--. q ~ ~~'~ 1~bryn>. Van1~~~0:~ ~-, <') Melliah.RJ' ;\,'f-~.j "'A"............ '~Sl_. 6Efate ...; ·;::tJ .... "'u "' u"' ;:s Combat Na1 ~ -· ~ naval operatioJ -N officers. ....... They "'0 ~ not only an ac ..8 charts and illus "0.... "'0 of these events . ..0.... ~ Material pr.i.n "'tJ -5 guarded ( r) in ~... ( 2) in avoiding :.a .... hearing of any < tJ "'c:: < CONFIDENTIAL CONFIDENTIAL NAVY DEPARTMENT Office of Naval Intelligence Washington, D. -
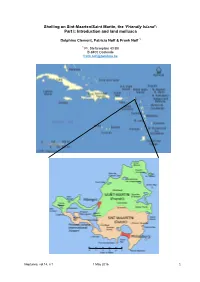
Shelling on Sint-Maarten/Saint Martin, the ‘Friendly Island ’: Part I: Introduction and Land Mollusca
Shelling on Sint-Maarten/Saint Martin, the ‘Friendly Island ’: Part I: Introduction and land mollusca Delphine Clement, Patricia Nolf & Frank Nolf 1 1 Pr. Stefanieplein 43 B8 B-8400 Oostende [email protected] Neptunea, vol.14, n°1 1 May 2016 1 Introduction Geography Sint-Maarten/Saint Martin (SXM) is situated at 18°04' N, 63°04′ W and belongs to what is called the 'Windward Islands', together with Puerto Rico, Saba, the Virgin Islands and some other islands, in contrast to the 'Leeward Islands' Aruba, Curaçao and Bonaire north of Venezuela. Saint Martin is confined by the Atlantic Ocean in the north and by the Caribbean Sea in the south. Saint Martin belongs to the northernmost islands of the Lesser Antilles in the external volcanic arc. It consists of two parts: the French Saint-Martin in the north and the Dutch Saint Maarten in the south. The total area is 86 km², of which about 34 km² is inhabited by approximately 35,000 of people in the Dutch Saint Martin. The ‘Collectivité de Saint Martin’, is an overseas collectivity of France and has an area of about 53 km², inhabited by more than 32,000 inhabitants. The capital of the Dutch part is Philipsburg while Marigot is the French capital. The island is very hilly and is dominated by several 'Hills' or 'Monts' such as the Mt. France, Mt. Boeuf, Mt. Caretta, Sentry Hill and above all the highest hill, the Pic Paradis (424 m). Ridges of the hills run into the sea and with a certain regularity around the coastline which is interrupted by numerous bays and white beaches, where hundreds of hotels and resorts shoot up like mushrooms. -

Deep-Water Photinae (Gastropoda: Nassariidae) from Eastern Africa, with Descriptions of Five New Species
European Journal of Taxonomy 720: 144–169 ISSN 2118-9773 https://doi.org/10.5852/ejt.2020.720.1123 www.europeanjournaloftaxonomy.eu 2020 · Fraussen K. et al. This work is licensed under a Creative Commons Attribution License (CC BY 4.0). Research article urn:lsid:zoobank.org:pub:C77C4C91-762D-4A06-82FA-3C58294E1570 Deep-water Photinae (Gastropoda: Nassariidae) from eastern Africa, with descriptions of five new species Koen FRAUSSEN 1, Lee Ann GALINDO 2 & José ROSADO 3 1,2 Research Associate Institute of Systematics, Evolution, Biodiversity (ISYEB), Muséum national d’histoire naturelle (MNHN), CNRS, SU, EPHE, UA, CP 51, Rue Cuvier 57, 75005 Paris, France. 1 Leuvensestraat 25, 3200 Aarschot, Belgium. 3 Avenida Friedrich Engels 373-1°, 1101 Maputo, Mozambique. 1 Corresponding author: [email protected] 2 Email: [email protected] 3 Email: [email protected] 1 urn:lsid:zoobank.org:author:5F9EFCF2-5BCF-486E-8D3E-088D85C45882 2 urn:lsid:zoobank.org:author:B84DC387-F1A5-4FE4-80F2-5C93E41CEC15 3 urn:lsid:zoobank.org:author:0415DDDF-2BC1-40CC-9DF6-06146A31C7AB Abstract. Deep-water species from the western Indian Ocean off the East African coast and Madagascar, belonging to the subfamily Photinae, are discussed and compared with species from the West Pacific. Phos elegantissimus Hayashi & Habe, 1965, P. hirasei Sowerby, 1913 and P. laevis Kuroda & Habe in Habe, 1961 are recorded from Mozambique and/or from Madagascar, hereby extending their known range considerably into the western Indian Ocean. The East African specimens formerly assigned to Phos roseatus Hinds, 1844 are found to differ from this West Pacific species. In total, five species are described as new: Phos ganii sp. -

Florida Keys Species List
FKNMS Species List A B C D E F G H I J K L M N O P Q R S T 1 Marine and Terrestrial Species of the Florida Keys 2 Phylum Subphylum Class Subclass Order Suborder Infraorder Superfamily Family Scientific Name Common Name Notes 3 1 Porifera (Sponges) Demospongia Dictyoceratida Spongiidae Euryspongia rosea species from G.P. Schmahl, BNP survey 4 2 Fasciospongia cerebriformis species from G.P. Schmahl, BNP survey 5 3 Hippospongia gossypina Velvet sponge 6 4 Hippospongia lachne Sheepswool sponge 7 5 Oligoceras violacea Tortugas survey, Wheaton list 8 6 Spongia barbara Yellow sponge 9 7 Spongia graminea Glove sponge 10 8 Spongia obscura Grass sponge 11 9 Spongia sterea Wire sponge 12 10 Irciniidae Ircinia campana Vase sponge 13 11 Ircinia felix Stinker sponge 14 12 Ircinia cf. Ramosa species from G.P. Schmahl, BNP survey 15 13 Ircinia strobilina Black-ball sponge 16 14 Smenospongia aurea species from G.P. Schmahl, BNP survey, Tortugas survey, Wheaton list 17 15 Thorecta horridus recorded from Keys by Wiedenmayer 18 16 Dendroceratida Dysideidae Dysidea etheria species from G.P. Schmahl, BNP survey; Tortugas survey, Wheaton list 19 17 Dysidea fragilis species from G.P. Schmahl, BNP survey; Tortugas survey, Wheaton list 20 18 Dysidea janiae species from G.P. Schmahl, BNP survey; Tortugas survey, Wheaton list 21 19 Dysidea variabilis species from G.P. Schmahl, BNP survey 22 20 Verongida Druinellidae Pseudoceratina crassa Branching tube sponge 23 21 Aplysinidae Aplysina archeri species from G.P. Schmahl, BNP survey 24 22 Aplysina cauliformis Row pore rope sponge 25 23 Aplysina fistularis Yellow tube sponge 26 24 Aplysina lacunosa 27 25 Verongula rigida Pitted sponge 28 26 Darwinellidae Aplysilla sulfurea species from G.P.