REVIEW ARTICLE Evolution of the Shoulder Girdle with Special Reference to the Problems of the Clavicle Abstract Persistent Probl
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

The Shoulder Girdle and Anterior Limb of Drepanosaurus Unguicaudatus
<oological Journal of the Linnean Socieg (1994), Ill: 247-264. With 12 figures The shoulder girdle and anterior limb of Drepanosaurus unguicaudatus (Reptilia, Neodiapsida) from the upper Triassic (Norian) Downloaded from https://academic.oup.com/zoolinnean/article/111/3/247/2691415 by guest on 27 September 2021 of Northern Italy SILVIO RENESTO Dipartimento di Scienze della Terra, Universita degli Studi, Via Mangiagalli 34, I-20133 Milano, Italy Received January 1993, accepted for publication February 1994 A reinvestigation of the osteology of the holotype of Drepanosaurus unguicaudatus Pinna, 1980 suggests that in earlier descriptions some osteological features were misinterpreted, owing to the crushing of the bones and because taphonomic aspects were not considered. The pattern of the shoulder girdle and fore-limb was misunderstood: the supposed interclavicle is in fact the right scapula, and the bones previously identified as coracoid and scapula belong to the anterior limb. The new reconstruction of the shoulder girdle, along with the morphology of the phalanges and caudal vertebrae, leads to a new hypothesis about the mode oflife of this reptile. Drepanosaurus was probably an arboreal reptile which used its enormous claws to scrape the bark from trees, perhaps in search of insects, just as the modern pigmy anteater (Cyclopes) does. Available diagnostic characters place Drepanosaurus within the Neodiapsida Benton, but it is impossible to ascribe this genus to one or other of the two major neodiapsid lineages, the Archosauromorpha and the Lepidosauromorpha. ADDITIONAL KEY WORDS:-Functional morphology - taxonomy - taphonomy - palaeoecology . CONTENTS Introduction ................... 247 Taphonomy ..... .............. 249 Systematic palaeontology . .............. 251 Genus Drepanosaurus Pinna, 1980 .............. 252 Drepannsaurus unguicaudatus Pinna, 1980. -

Early Tetrapod Relationships Revisited
Biol. Rev. (2003), 78, pp. 251–345. f Cambridge Philosophical Society 251 DOI: 10.1017/S1464793102006103 Printed in the United Kingdom Early tetrapod relationships revisited MARCELLO RUTA1*, MICHAEL I. COATES1 and DONALD L. J. QUICKE2 1 The Department of Organismal Biology and Anatomy, The University of Chicago, 1027 East 57th Street, Chicago, IL 60637-1508, USA ([email protected]; [email protected]) 2 Department of Biology, Imperial College at Silwood Park, Ascot, Berkshire SL57PY, UK and Department of Entomology, The Natural History Museum, Cromwell Road, London SW75BD, UK ([email protected]) (Received 29 November 2001; revised 28 August 2002; accepted 2 September 2002) ABSTRACT In an attempt to investigate differences between the most widely discussed hypotheses of early tetrapod relation- ships, we assembled a new data matrix including 90 taxa coded for 319 cranial and postcranial characters. We have incorporated, where possible, original observations of numerous taxa spread throughout the major tetrapod clades. A stem-based (total-group) definition of Tetrapoda is preferred over apomorphy- and node-based (crown-group) definitions. This definition is operational, since it is based on a formal character analysis. A PAUP* search using a recently implemented version of the parsimony ratchet method yields 64 shortest trees. Differ- ences between these trees concern: (1) the internal relationships of aı¨stopods, the three selected species of which form a trichotomy; (2) the internal relationships of embolomeres, with Archeria -
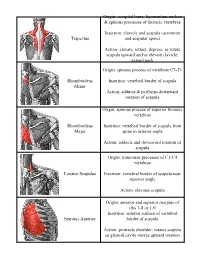
Trapezius Origin: Occipital Bone, Ligamentum Nuchae & Spinous Processes of Thoracic Vertebrae Insertion: Clavicle and Scapul
Origin: occipital bone, ligamentum nuchae & spinous processes of thoracic vertebrae Insertion: clavicle and scapula (acromion Trapezius and scapular spine) Action: elevate, retract, depress, or rotate scapula upward and/or elevate clavicle; extend neck Origin: spinous process of vertebrae C7-T1 Rhomboideus Insertion: vertebral border of scapula Minor Action: adducts & performs downward rotation of scapula Origin: spinous process of superior thoracic vertebrae Rhomboideus Insertion: vertebral border of scapula from Major spine to inferior angle Action: adducts and downward rotation of scapula Origin: transverse precesses of C1-C4 vertebrae Levator Scapulae Insertion: vertebral border of scapula near superior angle Action: elevates scapula Origin: anterior and superior margins of ribs 1-8 or 1-9 Insertion: anterior surface of vertebral Serratus Anterior border of scapula Action: protracts shoulder: rotates scapula so glenoid cavity moves upward rotation Origin: anterior surfaces and superior margins of ribs 3-5 Insertion: coracoid process of scapula Pectoralis Minor Action: depresses & protracts shoulder, rotates scapula (glenoid cavity rotates downward), elevates ribs Origin: supraspinous fossa of scapula Supraspinatus Insertion: greater tuberacle of humerus Action: abduction at the shoulder Origin: infraspinous fossa of scapula Infraspinatus Insertion: greater tubercle of humerus Action: lateral rotation at shoulder Origin: clavicle and scapula (acromion and adjacent scapular spine) Insertion: deltoid tuberosity of humerus Deltoid Action: -

Coracoid Process Anatomy: a Cadaveric Study of Surgically Relevant Structures Jorge Chahla, M.D., Ph.D., Daniel Cole Marchetti, B.A., Gilbert Moatshe, M.D., Márcio B
Quantitative Assessment of the Coracoacromial and the Coracoclavicular Ligaments With 3-Dimensional Mapping of the Coracoid Process Anatomy: A Cadaveric Study of Surgically Relevant Structures Jorge Chahla, M.D., Ph.D., Daniel Cole Marchetti, B.A., Gilbert Moatshe, M.D., Márcio B. Ferrari, M.D., George Sanchez, B.S., Alex W. Brady, M.Sc., Jonas Pogorzelski, M.D., M.H.B.A., George F. Lebus, M.D., Peter J. Millett, M.D., M.Sc., Robert F. LaPrade, M.D., Ph.D., and CAPT Matthew T. Provencher, M.D., M.C., U.S.N.R. Purpose: To perform a quantitative anatomic evaluation of the (1) coracoid process, specifically the attachment sites of the conjoint tendon, the pectoralis minor, the coracoacromial ligament (CAL), and the coracoclavicular (CC) ligaments in relation to pertinent osseous and soft tissue landmarks; (2) CC ligaments’ attachments on the clavicle; and (3) CAL attachment on the acromion in relation to surgically relevant anatomic landmarks to assist in planning of the Latarjet procedure, acromioclavicular (AC) joint reconstructions, and CAL resection distances avoiding iatrogenic injury to sur- rounding structures. Methods: Ten nonpaired fresh-frozen human cadaveric shoulders (mean age 52 years, range 33- 64 years) were included in this study. A 3-dimensional coordinate measuring device was used to quantify the location of pertinent bony landmarks and soft tissue attachment areas. The ligament and tendon attachment perimeters and center points on the coracoid, clavicle, and acromion were identified and subsequently dissected off the bone. Coordinates of points along the perimeters of attachment sites were used to calculate areas, whereas coordinates of center points were used to determine distances between surgically relevant attachment sites and pertinent bony landmarks. -

Anatomy and Relationships of the Triassic Temnospondyl Sclerothorax
Anatomy and relationships of the Triassic temnospondyl Sclerothorax RAINER R. SCHOCH, MICHAEL FASTNACHT, JÜRGEN FICHTER, and THOMAS KELLER Schoch, R.R., Fastnacht, M., Fichter, J., and Keller, T. 2007. Anatomy and relationships of the Triassic temnospondyl Sclerothorax. Acta Palaeontologica Polonica 52 (1): 117–136. Recently, new material of the peculiar tetrapod Sclerothorax hypselonotus from the Middle Buntsandstein (Olenekian) of north−central Germany has emerged that reveals the anatomy of the skull and anterior postcranial skeleton in detail. Despite differences in preservation, all previous plus the new finds of Sclerothorax are identified as belonging to the same taxon. Sclerothorax is characterized by various autapomorphies (subquadrangular skull being widest in snout region, ex− treme height of thoracal neural spines in mid−trunk region, rhomboidal interclavicle longer than skull). Despite its pecu− liar skull roof, the palate and mandible are consistent with those of capitosauroid stereospondyls in the presence of large muscular pockets on the basal plate, a flattened edentulous parasphenoid, a long basicranial suture, a large hamate process in the mandible, and a falciform crest in the occipital part of the cheek. In order to elucidate the phylogenetic position of Sclerothorax, we performed a cladistic analysis of 18 taxa and 70 characters from all parts of the skeleton. According to our results, Sclerothorax is nested well within the higher stereospondyls, forming the sister taxon of capitosauroids. Palaeobiologically, Sclerothorax is interesting for its several characters believed to correlate with a terrestrial life, although this is contrasted by the possession of well−established lateral line sulci. Key words: Sclerothorax, Temnospondyli, Stereospondyli, Buntsandstein, Triassic, Germany. -

A Unified Anatomy Ontology of the Vertebrate Skeletal System
A Unified Anatomy Ontology of the Vertebrate Skeletal System Wasila M. Dahdul1,2*, James P. Balhoff2,3, David C. Blackburn4, Alexander D. Diehl5, Melissa A. Haendel6, Brian K. Hall7, Hilmar Lapp2, John G. Lundberg8, Christopher J. Mungall9, Martin Ringwald10, Erik Segerdell6, Ceri E. Van Slyke11, Matthew K. Vickaryous12, Monte Westerfield11,13, Paula M. Mabee1 1 Department of Biology, University of South Dakota, Vermillion, South Dakota, United States of America, 2 National Evolutionary Synthesis Center, Durham, North Carolina, United States of America, 3 Department of Biology, University of North Carolina, Chapel Hill, North Carolina, United States of America, 4 Department of Vertebrate Zoology and Anthropology, California Academy of Sciences, San Francisco, California, United States of America, 5 The Jacobs Neurological Institute, University at Buffalo, Buffalo, New York, United States of America, 6 Oregon Health and Science University, Portland, Oregon, United States of America, 7 Department of Biology, Dalhousie University, Halifax, Nova Scotia, Canada, 8 Department of Ichthyology, The Academy of Natural Sciences, Philadelphia, Pennsylvania, United States of America, 9 Genomics Division, Lawrence Berkeley National Laboratory, Berkeley, California, United States of America, 10 The Jackson Laboratory, Bar Harbor, Maine, United States of America, 11 Zebrafish Information Network, University of Oregon, Eugene, Oregon, United States of America, 12 Department of Biomedical Sciences, Ontario Veterinary College, University of Guelph, Guelph, -

Reconstructing Pectoral Appendicular Muscle Anatomy in Fossil Fish and Tetrapods Over the Fins-To-Limbs Transition
Biol. Rev. (2017), pp. 000–000. 1 doi: 10.1111/brv.12386 Reconstructing pectoral appendicular muscle anatomy in fossil fish and tetrapods over the fins-to-limbs transition Julia L. Molnar1,∗ , Rui Diogo2, John R. Hutchinson3 and Stephanie E. Pierce4 1Department of Anatomy, New York Institute of Technology College of Osteopathic Medicine, Northern Boulevard, Old Westbury, NY, U.S.A. 2Department of Anatomy, Howard University College of Medicine, 520 W St. NW, Numa Adams Building, Washington, DC 20059, U.S.A. 3Structure and Motion Lab, Royal Veterinary College, Hawkshead Lane, Hatfield, Hertfordshire AL9 7TA, UK 4Museum of Comparative Zoology and Department of Organismic and Evolutionary Biology, Harvard University, 26 Oxford Street, Cambridge, MA 02138, U.S.A. ABSTRACT The question of how tetrapod limbs evolved from fins is one of the great puzzles of evolutionary biology. While palaeontologists, developmental biologists, and geneticists have made great strides in explaining the origin and early evolution of limb skeletal structures, that of the muscles remains largely unknown. The main reason is the lack of consensus about appendicular muscle homology between the closest living relatives of early tetrapods: lobe-finned fish and crown tetrapods. In the light of a recent study of these homologies, we re-examined osteological correlates of muscle attachment in the pectoral girdle, humerus, radius, and ulna of early tetrapods and their close relatives. Twenty-nine extinct and six extant sarcopterygians were included in a meta-analysis using information from the literature and from original specimens, when possible. We analysed these osteological correlates using parsimony-based character optimization in order to reconstruct muscle anatomy in ancestral lobe-finned fish, tetrapodomorph fish, stem tetrapods, and crown tetrapods. -

Breathing and Locomotion in Birds
Breathing and locomotion in birds. A thesis submitted to the University of Manchester for the degree of Doctor of Philosophy in the Faculty of Life Sciences. 2010 Peter George Tickle Contents Abstract 4 Declaration 5 Copyright Statement 6 Author Information 7 Acknowledgements 9 Organisation of this PhD thesis 10 Chapter 1 General Introduction 13 1. Introduction 14 1.1 The Avian Respiratory System 14 1.1.1 Structure of the lung and air sacs 16 1.1.2 Airflow in the avian respiratory system 21 1.1.3 The avian aspiration pump 25 1.2 The uncinate processes in birds 29 1.2.1 Uncinate process morphology and biomechanics 32 1.3 Constraints on breathing in birds 33 1.3.1 Development 33 1.3.2 Locomotion 35 1.3.2.1 The appendicular skeleton 35 1.3.2.2 Overcoming the trade-off between breathing 36 and locomotion 1.3.2.3 Energetics of locomotion in birds 38 1.4 Evolution of the ventilatory pump in birds 41 1.5 Overview and Thesis Aims 42 2 Chapter 2 Functional significance of the uncinate processes in birds. 44 Chapter 3 Ontogenetic development of the uncinate processes in the 45 domestic turkey (Meleagris gallopavo). Chapter 4 Uncinate process length in birds scales with resting metabolic rate. 46 Chapter 5 Load carrying during locomotion in the barnacle goose (Branta 47 leucopsis): The effect of load placement and size. Chapter 6 A continuum in ventilatory mechanics from early theropods to 48 extant birds. Chapter 7 General Discussion 49 References 64 3 Abstract of a thesis by Peter George Tickle submitted to the University of Manchester for the degree of PhD in the Faculty of Life Sciences and entitled ‘Breathing and Locomotion in Birds’. -

Comparative Bone Histology of the Turtle Shell (Carapace and Plastron)
Comparative bone histology of the turtle shell (carapace and plastron): implications for turtle systematics, functional morphology and turtle origins Dissertation zur Erlangung des Doktorgrades (Dr. rer. nat.) der Mathematisch-Naturwissenschaftlichen Fakultät der Rheinischen Friedrich-Wilhelms-Universität zu Bonn Vorgelegt von Dipl. Geol. Torsten Michael Scheyer aus Mannheim-Neckarau Bonn, 2007 Angefertigt mit Genehmigung der Mathematisch-Naturwissenschaftlichen Fakultät der Rheinischen Friedrich-Wilhelms-Universität Bonn 1 Referent: PD Dr. P. Martin Sander 2 Referent: Prof. Dr. Thomas Martin Tag der Promotion: 14. August 2007 Diese Dissertation ist 2007 auf dem Hochschulschriftenserver der ULB Bonn http://hss.ulb.uni-bonn.de/diss_online elektronisch publiziert. Rheinische Friedrich-Wilhelms-Universität Bonn, Januar 2007 Institut für Paläontologie Nussallee 8 53115 Bonn Dipl.-Geol. Torsten M. Scheyer Erklärung Hiermit erkläre ich an Eides statt, dass ich für meine Promotion keine anderen als die angegebenen Hilfsmittel benutzt habe, und dass die inhaltlich und wörtlich aus anderen Werken entnommenen Stellen und Zitate als solche gekennzeichnet sind. Torsten Scheyer Zusammenfassung—Die Knochenhistologie von Schildkrötenpanzern liefert wertvolle Ergebnisse zur Osteoderm- und Panzergenese, zur Rekonstruktion von fossilen Weichgeweben, zu phylogenetischen Hypothesen und zu funktionellen Aspekten des Schildkrötenpanzers, wobei Carapax und das Plastron generell ähnliche Ergebnisse zeigen. Neben intrinsischen, physiologischen Faktoren wird die -

Download PDF 3.83 MB
Evolutionary Development of the Mammalian Presternum Zeynep Metin Yozgyur Submitted in Partial Fulfillment of the Prerequisite for Honors in Biological Sciences under the advisement of Emily Buchholtz April 2019 This material is copyrighted by Zeynep Yozgyur and Emily Buchholtz, April 25, 2019. © 2019 Zeynep Yozgyur 1 Abstract The mammalian sternum has undergone a reduction in relative size and complexity over evolutionary time. This transformation was highly variable across species, generating multiple, controversial interpretations. Some authors claim that the evolutionary reduction led to loss of presternal elements, while others believe that all, or some, presternal elements were fused into a structure ambiguously referred to as the “manubrium”, a term adopted from the post- interclavicular unit of pre-mammalian ancestors. Previous work on the Paramylodon harlani presternum revealed a composite presternum and identified three elements - mediocranial, mediocaudal and lateral - each with a different developmental origin and marginal articulation. This project used medical and micro CT scans to determine if these elements are conserved across species and if there is a characteristic histology associated with each. All three elements were identified in humans based on their locations and articulations. Their fusion during ontogeny was documented. Elements could not be associated with a characteristic histology. Instead, histology appears to reflect the mechanical forces to which different regions of the presternum are subjected, whatever their developmental origin. This opens the possibility that the presternum can be used to infer the limb use and locomotor style of extinct taxa. These lines of evidence indicate that the reduction of relative sternal size seen over evolutionary time has not led to element loss. -

Some Notes on the Diverse Brachiosaurid Sauropods of the Late Jurassic of North America, Europe and Africa
Some Notes on the Diverse Brachiosaurid Sauropods of the Late Jurassic of North America, Europe and Africa GREGORY S. PAUL 3109 N. Calvert St. Baltimore, Maryland 21218 USA [email protected] June 2012 Abstract: The unusual placement of the parapophyses on elongotated peduncles further distinguishes Brachiosaurus from other brachiosaurid genera including Giraffatitan. Late Jurassic brachiosaurid taxonomy is probably more complex than has been realized, in part because of evolution over the considerable periods of times recorded in the Morrison and Tendaguru formations. The presence of reduced tails on late Jurassic brachiosaurids is confirmed. INTRODUCTION It was long assumed that the brachiosaurid material found in the Late Jurassic Morrison, Tendaguru, and Lourinha Formations from three continents belonged to the genus Brachiosaurus (as per Janensch 1950, 1961, Lapparent and Zbyszewski 1957, Jensen 1987) until Paul (1988) noted significant differences between the type species B. altithorax (Fig. 1A) from North America and the African B. brancai (Fig. 1B) indicated at least a subgeneric separation, and Taylor (2009) formally separated the sauropods, leaving much of the Tendaguru brachiosaurids material titled Giraffatitan brancai; the generic separation has been tentatively questioned (Chure et al. 2010). In addition European B. alatalaiensis has been retitled Lusotitan alatalaiensis (Antunes and Mateus 2003), and a dwarf brachiosaur from the European Mittlere Kimmeridge-Stufe deposits has been designated Europasaurus holgeri (Sander et al. 2006). Institutional Abbreviations: AMNH, American Museum of Natural History, New York; BYU, Brigham Young University, Provo; FMNH, Field Museum of Natural History, Chicago; HMN, Humboldt Museum fur Naturkunde, Berlin; NHMUK, Natural History Museum, London; USNM, United States Natural History Museum, Washington DC. -

Rehabilitation Guidelines for Latarjet/Coracoid Process Transfer Josef K
Rehabilitation Guidelines for Latarjet/Coracoid Process Transfer Josef K. Eichinger, MD Shoulder instability may be caused from congenital deformity, recurrent overuse activity, and/or traumatic dislocation. Surgical stabilization of the glenohumeral joint is indicated after conservative treatment fails and recurrent instability/subluxation continues. A number of different surgical procedures may be indicated in this situation, often divided into soft tissue or bony procedures. Shoulder Instability – Soft Tissue: Surgical reconstruction targeting the glenohumeral joint’s soft tissues for shoulder instability, typically involves labral repairs, the most common being the Bankart repair. A Bankart lesion typically occurs from an anterior-inferior dislocation of the humerus, tearing the labrum from it’s attachment to the glenoid, thereby detaching the inferior gleno-humeral ligament (IGHL). Surgical management of this revolves around labral repair to reattach the IGHL under appropriate tension. This may be accomplished either arthroscopically or through an open approach.1 Most traumatic glenohumeral dislocations may not only cause a Bankart lesion, but may create impression fractures in the postero-superior humeral head termed Hill-Sachs lesions.2 An adverse effect from this procedure includes suturing the capsule too tightly, causing a shortening of the capsule, and thus decreasing the external rotation allowed at the glenohumeral joint. Other complications are extremely rare, but may include axillary nerve damage, subscapularis rupture (seen only in open repairs performed with subscapularis detachment and repair), and recurrent instability. If there is bony deficiency in the glenoid, which represents 20% or more of the antero-inferior glenoid, it is biomechanically impossible to restore the same stability and is therefore more prone to recurrent instability and failure.