Historical Rates of Sediment and Nutrient Accumulation in Marshes of the Upper St
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Final Summary Document of Public Scoping Comments Submitted by the 3/31/11 Deadline
U.S. Fish and Wildlife Service Proposed Everglades Headwaters National Wildlife Refuge and Conservation Area Summary of Public Scoping Comments - as of 3.31.2011 Comments were submitted in a variety of ways (e.g., at a public scoping meeting and by mail, fax, and email). Attendance at the public scoping meetings averaged ~440 per meeting: ~200 in Sebring, ~325 in Kissimmee, ~665 in Okeechobee, and ~580 in Vero Beach. As of March 31, 2011, the deadline for public scoping comments, over 38,000 written comments had been received. The comments were summarized and are grouped together by topic, as listed. • Wildlife and Habitat • Resource Protection • Recreation • Administration • General/Other Comments List of acronyms used in comments: BLM Bureau of Land Management CERP Comprehensive Everglades Restoration Plan DEP Florida Department of Environmental Protection DOI U.S. Department of Interior DOT Florida Department of Transportation EH Everglades Headwaters ESV Ecosystem Services Values FDOT Florida Department of Transportation FWC Florida Fish and Wildlife Conservation Commission FWS U.S. Fish and Wildlife Service, also USFWS LOPP Lake Okeechobee Protection Plan NEPA National Environmental Policy Act NPS National Park Service NRC National Research Council NWR National Wildlife Refuge PES Payments for Ecosystem Services SFWMD South Florida Water Management District STA Stormwater Treatment Area TEV Total Economic Value TNC The Nature Conservancy USFWS U.S. Fish and Wildlife Service, also FWS Wildlife and Habitat General • If worried about the environment, we had more endangered species than anywhere in the State. We have the same amount of endangered species. We are good land stewards, so we don’t need the government or anything else. -

Professional Paper SJ98-PP1
Professional Paper SJ98-PP1 5ohns * A Professional Paper SJ98-PP1 THE UPPER ST. JOHNS RIVER BASIN PROJECT THE ENVIRONMENTAL TRANSFORMATION OF A PUBLIC FLOOD CONTROL PROJECT by Maurice Sterling Charles A. Padera St. Johns J^iver Water Management District Palatka, Florida 1998 Northwest Florida Water Management District anagement Southwest Florida Water Management District St. Johns River Water Management District South Florida Water Management District The St. Johns River Water Management District (SJRWMD) was created by the Florida Legislature in 1972 to be one of five water management districts in Florida. It includes all or part of 19 counties in northeast Florida. The mission of SJRWMD is to manage water resources to ensure their continued availability while maximizing both environmental and economic benefits. It accomplishes its mission through regulation; applied research; assistance to federal, state, and local governments; operation and maintenance of water control works; and land acquisition and management. Professional Papers are published to disseminate information collected by SJRWMD in pursuit of its mission. Copies of this report can be obtained from: Library St. Johns River Water Management District P.O. Box 1429 Palatka, FL 32178-1429 Phone: (904) 329-4132 Professional Paper SJ98-PP1 The Upper St. Johns River Basin Project The Environmental Transformation of a Public Flood Control Project Maurice Sterling and Charles A. Padera St. Johns River Water Management District, Palatka, Florida ABSTRACT During the 1980s, the Upper St. Johns River Basin Project was transformed into a national model of modern floodplain management. The project is sponsored jointly by the St. Johns River Water Management District and the U.S. -
Appendix 1 U.S
U.S. Department of the Interior Prepared in cooperation with the Appendix 1 U.S. Geological Survey Florida Department of Agriculture and Consumer Services, Office of Agricultural Water Policy Open-File Report 2014−1257 81°45' 81°30' 81°15' 81°00' 80°45' 524 Jim Creek 1 Lake Hart 501 520 LAKE 17 ORANGE 417 Lake Mary Jane Saint Johns River 192 Boggy Creek 535 Shingle Creek 519 429 Lake Preston 95 17 East Lake Tohopekaliga Saint Johns River 17 Reedy Creek 28°15' Lake Lizzie Lake Winder Saint Cloud Canal ! Lake Tohopekaliga Alligator Lake 4 Saint Johns River EXPLANATION Big Bend Swamp Brick Lake Generalized land use classifications 17 for study purposes: Crabgrass Creek Land irrigated Lake Russell Lake Mattie Lake Gentry Row crops Lake Washington Peppers−184 acresLake Lowery Lake Marion Creek 192 Potatoes−3,322 acres 27 Lake Van Cantaloupes−633 acres BREVARD Lake Alfred Eggplant−151 acres All others−57 acres Lake Henry ! UnverifiedLake Haines crops−33 acres Lake Marion Saint Johns River Jane Green Creek LakeFruit Rochelle crops Cypress Lake Blueberries−41 acres Citrus groves−10,861 acres OSCEOLA Peaches−67 acresLake Fannie Lake Hamilton Field Crops Saint Johns River Field corn−292 acres Hay−234 acres Lake Hatchineha Rye grass−477 acres Lake Howard Lake 17 Seeds−619 acres 28°00' Ornamentals and grasses Ornamentals−240 acres Tree nurseries−27 acres Lake Annie Sod farms−5,643Lake Eloise acres 17 Pasture (improved)−4,575 acres Catfish Creek Land not irrigated Abandoned groves−4,916 acres Pasture−259,823 acres Lake Rosalie Water source Groundwater−18,351 acres POLK Surface water−9,106 acres Lake Kissimmee Lake Jackson Water Management Districts irrigated land totals Weohyakapka Creek Tiger Lake South Florida Groundwater−18,351 acres 441 Surface water−7,596 acres Lake Marian St. -

Status of the Aquatic Plant Maintenance Program in Florida Public Waters
Status of the Aquatic Plant Maintenance Program in Florida Public Waters Annual Report for Fiscal Year 2006 - 2007 Executive Summary This report was prepared in accordance with §369.22 (7), Florida Statutes, to provide an annual assessment of the control achieved and funding necessary to manage nonindigenous aquatic plants in intercounty waters. The authority of the Department of Environmental Protection (DEP) as addressed in §369.20 (5), Florida Statutes, extends to the management of nuisance populations of all aquatic plants, both indigenous and nonindigenous, and in all waters accessible to the general public. The aquatic plant management program in Florida’s public waters involves complex operational and financial interactions between state, federal and local governments as well as private sector compa- nies. A summary of plant acres controlled in sovereignty public waters and associated expenditures contracted or monitored by the DEP during Fiscal Year 2006-2007 is presented in the tables on page 42 of this report. Florida’s aquatic plant management program mission is to reduce negative impacts from invasive nonindigenous plants like water hyacinth, water lettuce and hydrilla to conserve the multiple uses and functions of public lakes and rivers. Invasive plants infest 95 percent of the 437 public waters inventoried in 2007 that comprise 1.25 million acres of fresh water where fishing alone is valued at more than $1.5 billion annually. Once established, eradicating invasive plants is difficult or impossible and very expensive; therefore, continuous maintenance is critical to sustaining navigation, flood control and recreation while conserving native plant habitat on sovereignty state lands at the lowest feasible cost. -

2020 Integrated Water Quality Assessment for Florida: Sections 303(D), 305(B), and 314 Report and Listing Update
2020 Integrated Water Quality Assessment for Florida: Sections 303(d), 305(b), and 314 Report and Listing Update Division of Environmental Assessment and Restoration Florida Department of Environmental Protection June 2020 2600 Blair Stone Rd. Tallahassee, FL 32399-2400 floridadep.gov 2020 Integrated Water Quality Assessment for Florida, June 2020 This Page Intentionally Blank. Page 2 of 160 2020 Integrated Water Quality Assessment for Florida, June 2020 Letter to Floridians Ron DeSantis FLORIDA DEPARTMENT OF Governor Jeanette Nuñez Environmental Protection Lt. Governor Bob Martinez Center Noah Valenstein 2600 Blair Stone Road Secretary Tallahassee, FL 32399-2400 June 16, 2020 Dear Floridians: It is with great pleasure that we present to you the 2020 Integrated Water Quality Assessment for Florida. This report meets the Federal Clean Water Act reporting requirements; more importantly, it presents a comprehensive analysis of the quality of our waters. This report would not be possible without the monitoring efforts of organizations throughout the state, including state and local governments, universities, and volunteer groups who agree that our waters are a central part of our state’s culture, heritage, and way of life. In Florida, monitoring efforts at all levels result in substantially more monitoring stations and water quality data than most other states in the nation. These water quality data are used annually for the assessment of waterbody health by means of a comprehensive approach. Hundreds of assessments of individual waterbodies are conducted each year. Additionally, as part of this report, a statewide water quality condition is presented using an unbiased random monitoring design. These efforts allow us to understand the state’s water conditions, make decisions that further enhance our waterways, and focus our efforts on addressing problems. -

Technical Publication Sj 87-3 Establishment of Minimum Surface Water Requirements for the Greater Lake Washington Basin
TECHNICAL PUBLICATION SJ 87-3 ESTABLISHMENT OF MINIMUM SURFACE WATER REQUIREMENTS FOR THE GREATER LAKE WASHINGTON BASIN BY G. B. Hall Division of Environmental Sciences Department of Water Resources St. Johns River Water Management District Palatka, Florida February, 1987 Project No. 20 024 24 ACKNOWLEDGEMENTS Brad Purcell provided assistance in graphics; Marvin Williams, Chris Mayberry and Palmer Kinser assisted in the analysis of aerial photography and the digitization and plotting of maps; and Phyllis Howard and Sherry Dearth typed the report. Drafts of the manuscript were critically read by Barbara Vergara, Edgar Lowe, Charles Tai, Don Rao, and Robert Massarelli (SBWA, Melbourne). ii TABLE OF CONTENTS PAGE ACKNOWLEDGEMENTS i i LIST OF FIGURES V LIST OF TABLES viii EXECUTIVE SUMMARY 1 INTRODUCTION 5 Background and Scope 5 The S tudy Area 9 Land-Use and Regional Importance 9 Soils 11 Climate 14 Ecological Alteration of the Floodplain 18 Surface Water Hydrology 25 METHODOLOGY 33 Environmental Analyses and Minimum Level Criteria Development 33 Basin Morphometry 33 Floodplain Vegetation 34 Minimum Level Criteria Development 36 RESULTS 40 Floodplain Alteration 40 Basin Morphometry 40 Floodplain Vegetation 40 Minimum Level Criteria 49 Floodplain Vegetation and Soils 49 Water Quality 54 Transfer of Detrital Materials and Sediments 63 Fish and Wildlife Habitat 64 Recreation and Navigation 65 Aesthetic and Scenic Attributes 66 DISCUSSION 67 LITERATURE CITED 72 APPENDIX A APPENDIX B IV LIST OF FIGURES FIGURE PAGE 1 Map of the Upper St. Johns River Basin showing the study area 7 2 Map of the Greater Lake Washington Basin, 3 Soils of the Upper St. -

Your Guide to Eating Fish Caught in Florida
Fish Consumption Advisories are published periodically by the Your Guide State of Florida to alert consumers about the possibility of chemically contaminated fish in Florida waters. To Eating The advisories are meant to inform the public of potential health risks of specific fish species from specific Fish Caught water bodies. In Florida February 2019 Florida Department of Health Prepared in cooperation with the Florida Department of Environmental Protection and Agriculture and Consumer Services, and the Florida Fish and Wildlife Conservation Commission 2019 Florida Fish Advisories • Table 1: Eating Guidelines for Fresh Water Fish from Florida Waters (based on mercury levels) page 1-50 • Table 2: Eating Guidelines for Marine and Estuarine Fish from Florida Waters (based on mercury levels) page 51-52 • Table 3: Eating Guidelines for species from Florida Waters with Heavy Metals (other than mercury), Dioxin, Pesticides, Polychlorinated biphenyls (PCBs), or Saxitoxin Contamination page 53-54 Eating Fish is an important part of a healthy diet. Rich in vitamins and low in fat, fish contains protein we need for strong bodies. It is also an excellent source of nutrition for proper growth and development. In fact, the American Heart Association recommends that you eat two meals of fish or seafood every week. At the same time, most Florida seafood has low to medium levels of mercury. Depending on the age of the fish, the type of fish, and the condition of the water the fish lives in, the levels of mercury found in fish are different. While mercury in rivers, creeks, ponds, and lakes can build up in some fish to levels that can be harmful, most fish caught in Florida can be eaten without harm. -

Annual Report of Activities Conducted Under the Cooperative Aquatic Plant Control Program in Florida Public Waters for Fiscal Year 2012-2013
Annual Report of Activities Conducted under the Cooperative Aquatic Plant Control Program in Florida Public Waters for Fiscal Year 2012-2013 Florida Fish and Wildlife Conservation Commission Invasive Plant Management Section Submitted by: FL Fish and Wildlife Conservation Commission Invasive Plant Management Section 3900 Commonwealth Blvd. MS705 Tallahassee, FL 32399 Phone: 850-617-9420 Fax: 850-922-1249 Annual Report of Activities Conducted under the Cooperative Aquatic Plant Control Program in Florida Public Waters for Fiscal Year 2012-2013 This report was prepared in accordance with §369.22 (7), Florida Statutes, to provide an annual summary of plants treated and funding necessary to manage aquatic plants in public waters. The Cooperative Aquatic Plant Control Program administered by the Florida Fish and Wildlife Conservation Commission (FWC) in Florida’s public waters involves complex operational and financial interactions between state, federal and local governments as well as private sector companies. FWC’s aquatic plant management program mission is to reduce negative impacts from invasive nonindigenous plants like water hyacinth, water lettuce and hydrilla to conserve the multiple uses and functions of public lakes and rivers. Invasive plants infest 96% of Florida’s 451 public waters inventoried in 2013 that comprise 1.26 million acres of fresh water. Once established, eradicating invasive plants is difficult or impossible and very expensive; therefore, continuous maintenance is critical to keep invasive plants at low levels to -
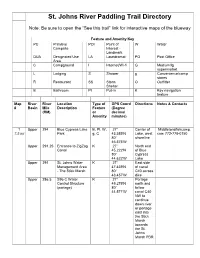
St. Johns River Blueway by Dean Campbell River Overview
St. Johns River Paddling Trail Directory Note: Be sure to open the “See this trail” link for interactive maps of the blueway Feature and Amenity Key PC Primitive POI Point of W Water Campsite Interest - Landmark DUA Designated Use LA Laundromat PO Post Office Area C Campground I Internet/Wi-fi G Medium/lg supermarket L Lodging S Shower g Convenience/camp stores R Restaurant SS Storm O Outfitter Shelter B Bathroom PI Put-in K Key navigation feature Map River River Location Type of GPS Coord Directions Notes & Contacts # Basin Mile Description Feature (Degree (RM) or decimal Amenity minutes) 1 Upper 294 Blue Cypress Lake B, PI, W, 27° Center of Middletonsfishcamp. 7.5 mi Park g, C 43.589'N Lake, west com 772-778-0150 80° shoreline 46.575'W Upper 291.25 Entrance to ZigZag K 27° North end Canal 45.222'N of Blue 80° Cypress 44.622'W Lake Upper 291 St. Johns Water K 27° East side Management Area 47.439'N of canal - The Stick Marsh 80° C40 across 43.457'W dike Upper 286.5 S96 C Water K 27° Portage Control Structure 49.279'N north and (portage) 80° follow 44.571'W canal C40 NW to continue down river or portage east into the Stick Marsh towards the St. Johns Marsh PBR Upper 286.5 St. Johns Marsh – B, PI, W 27° East side Barney Green 49.393'N of canal PBR* 80° C40 across 42.537'W dike 2 Upper 286.5 St. Johns Marsh – B, PI, W 27° East side 22 mi Barney Green 49.393'N of canal *2 PBR* 80° C40 across day 42.537'W dike trip Upper 279.5 Great Egret PC 27° East shore Campsite 54.627'N of canal 80° C40 46.177'W Upper 277 Canal Plug in C40 K 27° In canal -

Florida Fish and Wildlife Conservation Commission Statewide Alligator Harvest Data Summary
FWC Home : Wildlife & Habitats : Managed Species : Alligator Management Program FLORIDA FISH AND WILDLIFE CONSERVATION COMMISSION STATEWIDE ALLIGATOR HARVEST DATA SUMMARY YEAR AVERAGE LENGTH TOTAL HARVEST FEET INCHES 2000 8 8 2,552 2001 8 8.2 2,268 2002 8 3.7 2,164 2003 8 4.6 2,830 2004 8 5.8 3,237 2005 8 4.9 3,436 2006 8 4.8 6,430 2007 8 6.7 5,942 2008 8 5.1 6,204 2009 8 0 7,844 2010 7 10.9 7,654 2011 8 1.2 8,103 Provisional data 2000 STATEWIDE ALLIGATOR HARVEST DATA SUMMARY AVERAGE LENGTH TOTAL AREA NO AREA NAME FEET INCHES HARVEST 101 LAKE PIERCE 7 9.8 12 102 LAKE MARIAN 9 9.3 30 104 LAKE HATCHINEHA 8 7.9 36 105 KISSIMMEE RIVER (POOL A) 7 6.7 17 106 KISSIMMEE RIVER (POOL C) 8 8.3 17 109 LAKE ISTOKPOGA 8 0.5 116 110 LAKE KISSIMMEE 7 11.5 172 112 TENEROC FMA 8 6.0 1 402 EVERGLADES WMA (WCAs 2A & 2B) 8 8.2 12 404 EVERGLADES WMA (WCAs 3A & 3B) 8 10.4 63 405 HOLEY LAND WMA 9 11.0 2 500 BLUE CYPRESS LAKE 8 5.6 31 501 ST. JOHNS RIVER 1 8 2.2 69 502 ST. JOHNS RIVER 2 8 0.7 152 504 ST. JOHNS RIVER 4 8 3.6 83 505 LAKE HARNEY 7 8.7 65 506 ST. JOHNS RIVER 5 9 2.2 38 508 CRESCENT LAKE 8 9.9 23 510 LAKE JESUP 9 9.5 28 518 LAKE ROUSSEAU 7 9.3 32 520 LAKE TOHOPEKALIGA 9 7.1 47 547 GUANA RIVER WMA 9 4.6 5 548 OCALA WMA 9 8.7 4 549 THREE LAKES WMA 9 9.3 4 601 LAKE OKEECHOBEE (WEST) 8 11.7 448 602 LAKE OKEECHOBEE (NORTH) 9 1.8 163 603 LAKE OKEECHOBEE (EAST) 8 6.8 38 604 LAKE OKEECHOBEE (SOUTH) 8 5.2 323 711 LAKE HANCOCK 9 3.9 101 721 RODMAN RESERVOIR 8 7.0 118 722 ORANGE LAKE 8 9.3 125 723 LOCHLOOSA LAKE 9 3.4 56 734 LAKE SEMINOLE 9 1.5 16 741 LAKE TRAFFORD -
U N S U U S E U R a C S
Canaveral Natl VOLUSIA Seash Lake Dora Mount Dora O r a Midway Gopher n Wekiva River Slough g Lake Harris e Sanford B Heathrow l o Crystal o s d s Lake n o a m l John F Kennedy Lake Mary r Lake Dr Space Ctr O Tangerine T Harney 108th CongressLake Ola rl of the United States Little Lake Geneva Harris Mosquito Lagoon Lake Jessup StHwy 46 e Zellwood t Howey-in-the-Hills 6 Indian River R Astatula t 2 S 4 S tH DISTRICT 7 w Jordan Slough y 3 Turkey Lake Apopka Wekiwa Springs Longwood r ) John F Kennedy StHwy 414 a D ev Space Ctr Lake Brantley en Buck Lake (G SEMINOLE Mims 4 26 Rte St Winter Springs Puzzle Lake Max Hoeck 1 Creek StRd 46 (Main St) Forest City Casselberry Oviedo Bear Salt Lake StHwy 434 South Altamonte Fern Lake StHwyStHwy 406 StHwy 402 (Max Apopka Springs Park St Johns River StHwy 417 Chuluota 402 Brewer Memorial Pkwy) Canaveral Atlantic Ocean StHwy19 Lake Howell Mills Max Brewer Memorial Pkwy Lake Loughman Lake Natl Seash Lake Arthur Ferndale Maitland Schoolhouse Lake Lockhart Cape Rd Bear Gully Lake South StHwy 406 Lake Apopka Eaton- Clark Lake Lake Paradise Heights ville ( Garden St) W Shepherd a Fairview s Lake Lake h Banana Goldenrod i (Rouse Rd) (Rouse n Grassy DISTRICT Lake Shores 434 StHwy t) Creek Montverde Pickett S Lake Orlando Lake Osceola g th t u o 3 o n Mascotte Fox Lake S l ( A Pintail a ) v r 5 Creek t y e Lake n a 0 Lake 4 Lucy Cherry Lake Minneola Pine Winter Park e w Fairview C n y Lake ( e Sue 8 re w Hills Mills Ave 0 H StHwy 537 4 G t Lake ( 4 M DISTRICT S a y Titusville d Minneola a Lake w i r i n Starke -

Everglade Kite
EVERGLADENEWSLETTER OF THE KITE AUDUBON SOCIETY OF THE EVERGLADES Serving Palm Beach County, Florida Volume 54, Number 5 February 2014 CALENDAR Establishing Everglades Headwaters National Wildlife Refuge Complex Charlie Pelizza, Project Leader Sat. Feb 1, 7:45 a.m. STA 1E. ASE invites Tuesday, February 4, 7:30 p.m. Tropical Audubon to join us. Car pool around Storm Water Treatment Area managed by Charlie Pelizza is the Project Leader for the recently established Everglades Headwaters National South Florida Water Management District. Wildlife Refuge Complex. The Complex consists of Pelican Island Refuge, the Nation’s first Refuge Space limited. Call Linda to register: 561- established in 1903; Archie Carr, the most important stretch of nesting beach for loggerhead 742-7791 turtles in the western hemisphere and second in the world; Lake Wales Ridge, dominated by over Sun. Feb 2, 8:00 a.m. Pond Hawk Natural Area, 30 endemic and endangered scrub-dependent plants and animals; and Everglades Headwaters, the Boca Raton. 1501 Spanish River Blvd. Meet at 556th and one of the most recently established refuges. He received his bachelor’s degree at Elmira Library parking lot. Leader: Ben Kolstad College, and his Masters at the University of South Dakota. He lives in Wellington, Florida with Wed. Feb 5, 7:30 a.m. ARM Loxahatchee Sylvia, also a Project Leader with the Service, and two teenage children, Sierra and Kai. NWR. 10216 Lee Rd, Boynton Bch. Meet at Marsh Trail. Entry $5 per car Charlie has worked with Service, primarily in the Division of Refuges for 33 years in habitats Thur.