Estuary and Gulf of St. Lawrence Ecozone, SCCS, Dufour and Al
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Taking the Guesswork out of Lightning-Caused Wildfire Marjie Brown US Forest Service, [email protected]
University of Nebraska - Lincoln DigitalCommons@University of Nebraska - Lincoln JFSP Briefs U.S. Joint Fire Science Program 2008 Taking the Guesswork Out of Lightning-caused Wildfire Marjie Brown US Forest Service, [email protected] Follow this and additional works at: http://digitalcommons.unl.edu/jfspbriefs Part of the Forest Biology Commons, Forest Management Commons, Other Forestry and Forest Sciences Commons, and the Wood Science and Pulp, Paper Technology Commons Brown, Marjie, "Taking the Guesswork Out of Lightning-caused Wildfire" (2008). JFSP Briefs. 23. http://digitalcommons.unl.edu/jfspbriefs/23 This Article is brought to you for free and open access by the U.S. Joint Fire Science Program at DigitalCommons@University of Nebraska - Lincoln. It has been accepted for inclusion in JFSP Briefs by an authorized administrator of DigitalCommons@University of Nebraska - Lincoln. Lightning and fi re smoke. Taking the Guesswork Out of Lightning-caused Wildfi re Summary Lightning is a natural source of wildfi re ignitions and causes a substantial portion of large wildfi res across the globe. Simple predictions of lightning activity don’t accurately determine fi re ignition potential because fuel conditions must be considered in addition to the fact that most lightning is accompanied by signifi cant rain. Fire operations managers need improved tools for prediction of widespread dry thunderstorms, which are those that occur without signifi cant rainfall reaching the ground. It is these dry storms that generate lightning most likely to result in multiple fi re ignitions, often in remote areas. In previous work the researchers developed a formula that estimates the potential for cloud- to-ground lightning when dry thunderstorms are expected. -

Analysis of `Miholjday Summer' for Belgrade and Serbia Region
INTERNATIONAL JOURNAL OF CLIMATOLOGY Int. J. Climatol. 26: 1489–1499 (2006) Published online in Wiley InterScience (www.interscience.wiley.com) DOI: 10.1002/joc.1390 ANALYSIS OF ‘MIHOLJDAY SUMMER’ FOR BELGRADE AND SERBIA REGION NEDELJKO TODOROVIC´ a* and DRAGANA VUJOVIC´ b* a Hydrometeorological Service of Serbia, Belgrade, Yugoslavia b Department of Meteorology, Faculty of Physics, Belgrade, Yugoslavia Received 30 May 2006 Revised 14 February 2006 Accepted 11 April 2006 ABSTRACT Typical weather conditions with dry and warm features occur in autumn months, with temperatures above the normal temperatures for this period of the year for the Belgrade and Serbia region. Temperatures have values like the ones for the end of summer. That period of fair weather is called Miholjday (St Michael) summer (MS). An analysis of temperature has been the most important criterion for defining MS. Synoptic situation and temperature conditions during that period are analyzed and typical and atypical MSs are defined for Belgrade and Serbia region. The frequency of MS in the period 1946–2004 for Belgrade region is also analyzed. The general definition (Glossary of Meteorology) is assumed and we gave the specific definition of MS for Belgrade and Serbia region on the basis of real weather for longer series of observations. Copyright 2006 Royal Meteorological Society. KEY WORDS: summer climate variability; weather singularity; weather classification; synoptic climatology 1. INTRODUCTION Miholjday summer (MS) appears in autumn, in the period from the middle of September to the beginning of November. The name is related with Miholjday, the Christian feast celebrated on 12 October in the Orthodox Church (29 September in the Catholic Church). -
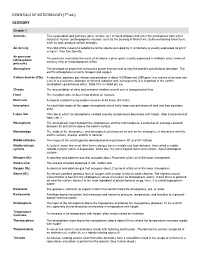
ESSENTIALS of METEOROLOGY (7Th Ed.) GLOSSARY
ESSENTIALS OF METEOROLOGY (7th ed.) GLOSSARY Chapter 1 Aerosols Tiny suspended solid particles (dust, smoke, etc.) or liquid droplets that enter the atmosphere from either natural or human (anthropogenic) sources, such as the burning of fossil fuels. Sulfur-containing fossil fuels, such as coal, produce sulfate aerosols. Air density The ratio of the mass of a substance to the volume occupied by it. Air density is usually expressed as g/cm3 or kg/m3. Also See Density. Air pressure The pressure exerted by the mass of air above a given point, usually expressed in millibars (mb), inches of (atmospheric mercury (Hg) or in hectopascals (hPa). pressure) Atmosphere The envelope of gases that surround a planet and are held to it by the planet's gravitational attraction. The earth's atmosphere is mainly nitrogen and oxygen. Carbon dioxide (CO2) A colorless, odorless gas whose concentration is about 0.039 percent (390 ppm) in a volume of air near sea level. It is a selective absorber of infrared radiation and, consequently, it is important in the earth's atmospheric greenhouse effect. Solid CO2 is called dry ice. Climate The accumulation of daily and seasonal weather events over a long period of time. Front The transition zone between two distinct air masses. Hurricane A tropical cyclone having winds in excess of 64 knots (74 mi/hr). Ionosphere An electrified region of the upper atmosphere where fairly large concentrations of ions and free electrons exist. Lapse rate The rate at which an atmospheric variable (usually temperature) decreases with height. (See Environmental lapse rate.) Mesosphere The atmospheric layer between the stratosphere and the thermosphere. -

Episodic Dust Events of Utah's Wasatch Front and Adjoining Region
1654 JOURNAL OF APPLIED METEOROLOGY AND CLIMATOLOGY VOLUME 51 Episodic Dust Events of Utah’s Wasatch Front and Adjoining Region W. JAMES STEENBURGH AND JEFFREY D. MASSEY Department of Atmospheric Sciences, University of Utah, Salt Lake City, Utah THOMAS H. PAINTER Jet Propulsion Laboratory, Pasadena, California (Manuscript received 4 January 2012, in final form 4 April 2012) ABSTRACT Episodic dust events cause hazardous air quality along Utah’s Wasatch Front and dust loading of the snowpack in the adjacent Wasatch Mountains. This paper presents a climatology of episodic dust events of the Wasatch Front and adjoining region that is based on surface weather observations from the Salt Lake City International Airport (KSLC), Geostationary Operational Environmental Satellite (GOES) imag- ery, and additional meteorological datasets. Dust events at KSLC—defined as any day [mountain standard time (MST)] with at least one report of a dust storm, blowing dust, and/or dust in suspension with a visi- bility of 10 km or less—average 4.3 per water year (WY: October–September), with considerable in- terannual variability and a general decline in frequency during the 1930–2010 observational record. The distributions of monthly dust-event frequency and total dust flux are bimodal, with primary and secondary maxima in April and September, respectively. Dust reports are most common in the late afternoon and evening. An analysis of the 33 most recent (2001–10 WY) events at KSLC indicates that 11 were associated with airmass convection, 16 were associated with a cold front or baroclinic trough entering Utah from the west or northwest, 4 were associated with a stationaryorslowlymovingfrontorbaroclinictroughwestof Utah, and 2 were associated with other synoptic patterns. -

Emerging Fashion Trends for Autumn Winter Season
IOSR Journal of Business and Management (IOSR-JBM) e-ISSN: 2278-487X, p-ISSN: 2319-7668. Volume 16, Issue 2. Ver. II (Feb. 2014), PP 46-47 www.iosrjournals.org Emerging Fashion Trends for Autumn Winter Season Muhukumar Nair* [Director-Vidya Institute of Fashion Technology, U.P. Technical University, Lucknow, India ] Abstract: This study was describe the values of trends and its need for the designers. Another purpose of the study to identify the duration of demands for the particular design/style. Thus the market value automatically high during the trend season and the merchandising process would across the break even point. Keywords : Color, Material, Market, Season, Stipulation. I. Introduction It's time to wrap up warm for autumn and winter, but there's no need to stop looking fabulous. Fashion trends this winter will let you retain a bit of glam - while also staying warm - and with versatility there's something to suit all styles and body shapes. Fashion trends change frequently keeping retail professionals on their toes. One of the most anticipated fashion trends involves the most popular colors which shift from season to season. Each year Pantone announces its top color often inspiring clothing and accessory designs for winter season, Pantone's top color is emerald green, and the color showed up consistently on the fall and winter runway shows indicating that it will be a mainstay of winter fashion. “During the cold and dark of Arctic winter, sea ice refreezes and achieves its maximum extent, usually in late Feb. or early Mar. According to a NASA analysis, this year the annual maximum extent was reached on Feb. -

An American Haboob U.S
S. B. Idso, R. S. Ingram, and J. M. Pritchard an american haboob U.S. Water Conservation Laboratory and National Weather Service, Phoenix, Ariz. 85040 1. Introduction by thunder and/or rain after a time interval varying up One of the world's most awesome displays of blowing to 2 hr. dust and sand is the legendary "haboob" of Khartoum in Satellite photographs have shown that the squall line the Sudan. Raised by strong winds often generated storms that develop south and east of Tucson appear by the outflow of rain-cooled air from a cumulonimbus to originate from large dense masses of clouds over the cloud, the leading edges of these storms take on the Sierra Madre Occidental of northern Sonora, Mexico. appearance of "solid walls" of dust that conform to These cloud masses over Sonora seem to form some the shape of a density current head and rise on the days rather explosively in the deep semi-tropical air average to between 1000 and 2000 meters (Sutton, 1925; mass. During this season of the year, the Bermuda High Lawson, 1971). The average speed of advance of the often extends westward into eastern Arizona, and during Khartoum haboobs is 32 mph, with the greatest speed the afternoon some of the Mexican activity has been seen being about 45 mph (Sutton, 1925). Maximum duration to move northwestward, steered by variations in the approaches 6-1/2 hr with a peak between 30 min and easterly flow (possibly easterly waves) on the bottom side one hour, the average duration being about 3 hr (Free- of the lobe of the Bermuda High. -

DUST DEVIL METEOROLOGY Jack R
NOAA~NWS /1(}q, 3~;).. ~ . NOAA Technicai ·Memoran_dlim Nws·CR-42 U.S. DEPARTMENT OF COMMERCE National Oceanic and Atmospheric Adminlstretian National Weather Service DUST 1DEV-IL... ,. METEOROLOGY.. .. ... .... - 1!. Jack ,R. .Cooley \ i ,. CiHTRAL REGION Kansas City.. Mo. () U. S. DEPARTMENT OF COMMERCE NATIONAL OCEANIC AND ATMOSPHERIC ADMINISTRATION NATIONAL WEATHER SERVICE NOAA Technical Memoran:l.um NWS CR-42 Il.JST DEVIL METEOROLOGY Jack R. Cooley CENTRAL REJJION KANSAS CITY, MISSOORI May 1971 CONTENTS () l. Page No. INTRODUCTION 1 1.1 Some Early Accounts of Dust Devils 1 1.2 Definition 2 1.3 The Atmospheric Circulation Family 2 1.4 Other Similar Small-Scale Circulations 2 1.41 Small Waterspouts 3 l. 42 Whirlies 3 1.43 Fire 1-!hirhdnds 3 1.44 Whirlwinds Associated With Cold Fronts 4 l. 5 Damage Caused by Dust Devils 4 1.51 Dust DeVils Vs. Aircraft Safety 5 2. FORMATION 5 2.1 Optimum Locations (Macro and Micro) 5 2.2 Optimum Time of Occurrence (During Day and Year) 6 2.3 Conditions Favoring Dust Devil Formation 7 2.31 Factors Favoring Steep Lapse Rates Near the Ground 7 A. Large Incident Solar Radiation Angles 7 B. Minimum Cloudiness 7 C. Lmr Humidity 8 D. Dry Barren Soil 8 E. Surface Winds Below a Critical Value 9 2.32 Potential Lapse Rates Near the Ground 9 2.33 Favorable Air Flow 12 2 ,34 Abundant Surface Material 12 2.35 Level Terrain 13 2.4 Triggering Devices 13 2.5 Size and Shape 14. 2.6 Dust Size·and Distribution 14 2.7 Duration 15 2.8 Direction of Rotation 16 2.9 Lateral Speed and Direction of Movement 17 3. -

Thunderstorm Terminology
Thunderstorm Terminology Thunderstorm — Updraft — Downdraft Cold front — Cumulonimbus cloud Latent heat — Isolated thunderstorms Cloudburst — Precipitation — Hail Thunderstorm lightning, thunder, rain, and dense clouds; may include heavy rain, hail, and strong winds Updraft an upward flow of air Downdraft a downward flow of air Cold front leading edge of a mass of heavy, colder air that is advancing Cumulonimbus giant clouds piled on top of each other; top cloud spreads out in the shape of an anvil heat released when water vapor condenses; is Latent heat the main driving energy of thunderstorms Isolated short-lived storms with light winds; usually do thunderstorms not produce violent weather on the ground Cloudburst sudden and short-lived heavy rainfall that usually occurs in a small area any moisture that falls from clouds and Precipitation reaches the ground Hail balls of ice about .2 inch (5 mm) to 6 inches (15 cm) that sometimes accompany storms WriteBonnieRose.com 1 Thunderstorm Terminology Dry thunderstorm — Thunder — Lightning Bead lightning — Ball lightning — Microburst Multiple-cell storms — Supercell Mesocyclone — Tornado Dry thunderstorm a storm where all raindrops evaporate while falling and none reach the ground sound heard when lightning heats the air so Thunder strongly and quickly it produces shock waves Lightning discharge of electricity after electric charges on particles in clouds grow large enough Bead lightning infrequent; looks like a string of bright spots; also called chain lightning Ball lightning rare; looks -

TREATISE ONLINE Number 48
TREATISE ONLINE Number 48 Part N, Revised, Volume 1, Chapter 31: Illustrated Glossary of the Bivalvia Joseph G. Carter, Peter J. Harries, Nikolaus Malchus, André F. Sartori, Laurie C. Anderson, Rüdiger Bieler, Arthur E. Bogan, Eugene V. Coan, John C. W. Cope, Simon M. Cragg, José R. García-March, Jørgen Hylleberg, Patricia Kelley, Karl Kleemann, Jiří Kříž, Christopher McRoberts, Paula M. Mikkelsen, John Pojeta, Jr., Peter W. Skelton, Ilya Tëmkin, Thomas Yancey, and Alexandra Zieritz 2012 Lawrence, Kansas, USA ISSN 2153-4012 (online) paleo.ku.edu/treatiseonline PART N, REVISED, VOLUME 1, CHAPTER 31: ILLUSTRATED GLOSSARY OF THE BIVALVIA JOSEPH G. CARTER,1 PETER J. HARRIES,2 NIKOLAUS MALCHUS,3 ANDRÉ F. SARTORI,4 LAURIE C. ANDERSON,5 RÜDIGER BIELER,6 ARTHUR E. BOGAN,7 EUGENE V. COAN,8 JOHN C. W. COPE,9 SIMON M. CRAgg,10 JOSÉ R. GARCÍA-MARCH,11 JØRGEN HYLLEBERG,12 PATRICIA KELLEY,13 KARL KLEEMAnn,14 JIřÍ KřÍž,15 CHRISTOPHER MCROBERTS,16 PAULA M. MIKKELSEN,17 JOHN POJETA, JR.,18 PETER W. SKELTON,19 ILYA TËMKIN,20 THOMAS YAncEY,21 and ALEXANDRA ZIERITZ22 [1University of North Carolina, Chapel Hill, USA, [email protected]; 2University of South Florida, Tampa, USA, [email protected], [email protected]; 3Institut Català de Paleontologia (ICP), Catalunya, Spain, [email protected], [email protected]; 4Field Museum of Natural History, Chicago, USA, [email protected]; 5South Dakota School of Mines and Technology, Rapid City, [email protected]; 6Field Museum of Natural History, Chicago, USA, [email protected]; 7North -

Autumn, the Neglected Season in Climate Change Research
TREE-1899; No. of Pages 8 Review Autumn, the neglected season in climate change research 1 1 2 Amanda S. Gallinat , Richard B. Primack , and David L. Wagner 1 Department of Biology, Boston University, 5 Cummington Mall, Boston, MA 02215, USA 2 Department of Ecology and Evolutionary Biology, University of Connecticut, 75 North Eagleville Road, U-43 Storrs, CT 06269, USA Autumn remains a relatively neglected season in climate identify five areas ripe for future research, and provide change research in temperate and arctic ecosystems. recommendations for research in those areas. This neglect occurs despite the importance of autumn events, including leaf senescence, fruit ripening, bird and What we know about autumn insect migration, and induction of hibernation and dia- In temperate ecosystems, the autumn phenomena that have pause. Changes in autumn phenology alter the repro- received the most attention in climate change research are ductive capacity of individuals, exacerbate invasions, leaf senescence and migratory bird departures. Insect allow pathogen amplification and higher disease-trans- diapause and fruit ripening have also garnered moderate mission rates, reshuffle natural enemy–prey dynamics, interest. Other autumn phenomena such as amphibian shift the ecological dynamics among interacting species, dormancy and bud formation remain less studied and poorly and affect the net productivity of ecosystems. We syn- understood. thesize some of our existing understanding of autumn Despite the relative neglect of autumn, ecologists have phenology and identify five areas ripe for future climate made important progress in understanding the drivers of change research. We provide recommendations to ad- autumn phenology and the effects of climate change on dress common pitfalls in autumnal research as well as to autumn events (Figure 1). -

Spring Winter Summer Autumn
• Always drive on good, properly inflated issouri is a state of four seasons tires. and each season has its own unique road conditions. Missouri driving • Know and obey all traffic laws. cannot be categorized entirely into spring, summer, autumn, or winter. Nature some- • Be ready to adjust your speed to be ap- times mixes our four seasons together, and propriate for constantly changing driving this can cause problems when we travel. conditions. This brochure has been prepared to give you some tips on how to handle our Finally, let’s all work together, so fewer many varied driving conditions. people will become traffic crash statistics on Missouri’s highways. Spring Buckle Up Missouri! • Never drive when you have been drink- ing alcoholic beverages. Summer • Never ride with someone who has been drinking. • If medication directions indicate you should not drive after taking it, don’t do Feel free to call the it. Road Condition Report Hotline at: Produced by: Public Information and Education Division • Have a good attitude when you drive. Be Published by: Autumn patient with others. 1-888-275-6636 Missouri State Highway Patrol 1510 East Elm Street • Give driving your full attention. Behind Or, check the Patrol’s Jefferson City, MO 65101 the wheel is no place to read, put on 573-751-3313 makeup, or talk on the cell phone. web site at: V/TDD 573-751-3313 email: [email protected] • How about those eyes? Don’t be vain. If www.mshp.dps.mo.gov www.mshp.dps.mo.gov Winter you need glasses, wear them. -

P1.4 West Texas Mesonet Observations of Wake Lows and Heat Bursts Across Northwest Texas
P1.4 WEST TEXAS MESONET OBSERVATIONS OF WAKE LOWS AND HEAT BURSTS ACROSS NORTHWEST TEXAS Mark R. Conder*, Steven R. Cobb, and Gary D. Skwira National Weather Service Forecast Office, Lubbock, Texas 1. INTRODUCTION Wake lows (WLs) and heat bursts (HBs) are mesoscale phenomena associated with thunderstorms that can result in strong and sometimes severe (>25.5 m s-1), near-surface winds. The operational detection of severe winds is problematic because the associated convection can appear quite innocuous via WSR-88D data. The thermodynamic environment that supports WL and HB development is similar, and is often compared to the classic “onion” sounding structure as described by Zipser (1977). Observational studies such as Johnson et al. (1989) show that nearly dry-adiabatic lapse rates are prevalent in the lower to mid-troposphere, with a shallow stable layer near ground level. This type of environment is most commonly observed in the high plains during the warm season. While documented heat bursts were infrequent in the past, the recent expansion of surface mesonetworks are increasing the likelihood that these events will be sampled. One such network is the West Texas Mesonet (WTXM), a collection of over 40 meteorological stations spread across the Panhandle and South Plains of northwest Texas. During the period 1 June 2004 to 30 August 2006, the authors have FIG. 1. The Study area including the WTXM station recorded 10 WL/HB events that have been sampled by locations. Not all stations were available for every stations of the WTXM. Some of the more extreme event. measurements include a 15 degrees Celsius increase in temperature at the Pampa and McClean WTXM sites in -1 Meteorological data from each WTXM station is one event and 35 m s wind gusts at sites in Brownfield recorded in 5-minute intervals, representing average and Jayton on separate occasions.