Cettia Diphone)
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Natural History of Japanese Birds
Natural History of Japanese Birds Hiroyoshi Higuchi English text translated by Reiko Kurosawa HEIBONSHA 1 Copyright © 2014 by Hiroyoshi Higuchi, Reiko Kurosawa Typeset and designed by: Washisu Design Office Printed in Japan Heibonsha Limited, Publishers 3-29 Kanda Jimbocho, Chiyoda-ku Tokyo 101-0051 Japan All rights reserved. No part of this publication may be reproduced or transmitted in any form or by any means without permission in writing from the publisher. The English text can be downloaded from the following website for free. http://www.heibonsha.co.jp/ 2 CONTENTS Chapter 1 The natural environment and birds of Japan 6 Chapter 2 Representative birds of Japan 11 Chapter 3 Abundant varieties of forest birds and water birds 13 Chapter 4 Four seasons of the satoyama 17 Chapter 5 Active life of urban birds 20 Chapter 6 Interesting ecological behavior of birds 24 Chapter 7 Bird migration — from where to where 28 Chapter 8 The present state of Japanese birds and their future 34 3 Natural History of Japanese Birds Preface [BOOK p.3] Japan is a beautiful country. The hills and dales are covered “satoyama”. When horsetail shoots come out and violets and with rich forest green, the river waters run clear and the moun- cherry blossoms bloom in spring, birds begin to sing and get tain ranges in the distance look hazy purple, which perfectly ready for reproduction. Summer visitors also start arriving in fits a Japanese expression of “Sanshi-suimei (purple mountains Japan one after another from the tropical regions to brighten and clear waters)”, describing great natural beauty. -

JAPANESE BUSH-WARBLER Cettia Diphone
JAPANESE BUSH-WARBLER Cettia diphone Other: Bush Warbler, Uguisu C. d. cantans? naturalized (non-native) resident, long established The Japanese Bush Warbler is native to Japan and surrounding islands, with northern populations being slightly migratory (AOU 1998). It and the Chinese or Manchurian bush-warbler (C. canturians) of e. China are closely related and sometimes considered conspecific (as "Bush Warbler"). Japanese Bush-Warblers have not been introduced anywhere in the world except the Southeastern Hawaiian Islands, where they were released on O'ahu in 1929-1941 (Caum 1933, Long 1981, Lever 1987) and have since spread naturally to most or all other Southeastern Islands. Concerns have been expressed about competition of bush-warblers for food with native species (Foster 2009). Japanese Bush-Warblers were initially introduced by the HBAF in 1929 to control insects, but several other releases on O'ahu (totaling approximately 138 individuals) were made by the Honolulu Mejiro Club and Hui Manu Society for aesthetic purposes, primarily or entirely in Nu'uanu Valley in 1931-1941 (Caum 1933; HAS 1967; Swedberg 1967a; Berger 1972, 1975c, 1981; E 17:2-3, 37:148; PoP 49[12]:29). They spread quickly on O'ahu, were noted by Munro (1944) in the Waianae Range by 1935, were considered established by Bryan (1941), were found commonly throughout both this and the Ko'olau Range by the 1950s (Northwood 1940, Pedley 1949; E 1[12]:3-4, 17:2-3, 25:91, 27:15-16, 31:108; summarized by Berger 1975c, Shallenberger 1977c, Shallenberger and Vaughn 1978), and were observed as far as Kahuku by 1977 (E 38:56). -

Download Okhotsk Checklist (105 KB)
Okhotsk Checklist Okhotsk Chapter of the Wild Bird Society of Japan Home Events News Checklist Bird Guide Birding Spots Surveys References Links About Us Contact Us -- Checklist of Birds in the Okhotsk Region Exactly 354 species of wild birds have been positively recorded thus far (as of 2 April 2013) in the Okhotsk (Sub-prefectural) region of Hokkaido. This compares with 633 species (excluding 43 introduced species) for all of Japan (Ornithological Society of Japan, 2012). It is a surprisingly large number in a region that has relatively few birdwatchers. The Okhotsk region is a wonderful field blessed with a great diversity of habitats. The checklist below contains all 354 of these species, plus 4 introduced species. If you happen to be in this region and spot any birds that are not in this list, please by all means report it to us! (via the Contact Us page). One point to note in this list, however, is that while we generally have one name in Japanese for our birds, in English there is often more than one name. There is also occasionally some disagreement in the references concerning scientific names. (Avibase is a good online source to check for updates to names in current use.) For now, we have included most of the recent names that we have come across for the benefit of our readers around the world. We will continue to edit this list over time based on new information as we receive it. To download a PDF version of the checklist page, click here → Download Okhotsk Checklist (105 KB). -

Differences in Vocalizations of Japanese Bush Warblers on Chichijima and Hahajima in the Ogasawara Islands
Bull. Natl. Mus. Nat. Sci., Ser. A, 44(4), pp. 125–132, November 22, 2018 Differences in Vocalizations of Japanese Bush Warblers on Chichijima and Hahajima in the Ogasawara Islands Shoji Hamao Department of Zoology, National Museum of Nature and Science, 4–1–1 Amakubo, Tsukuba, Ibaraki, 305–0005 Japan E-mail: [email protected] (Received 4 August 2018; accepted 26 September 2018) Abstract Bird songs play an important role in species recognition in reproductive behaviors. Therefore, geographic variation in the songs of a given species affects gene flow and reproductive isolation. A previous study showed that Japanese bush warblers, Cettia diphone, on Hahajima in the Ogasawara (Bonin) Islands sing songs with a simpler acoustic structure than those of C. diphone in mainland Japan. I investigated whether the acoustic structure of songs differs between the populations on two neighboring islands, Chichijima and Hahajima. In type-H songs, the maxi- mum frequency was lower, the minimum frequency was higher, and the frequency range was, therefore, narrower on Hahajima than on Chichijima. In type-L songs, the maximum and minimum frequencies were lower and the frequency range was narrower on Hahajima than on Chichijima. A possible factor giving rise to the narrower frequency range on Hahajima is the sound transmission properties of the habitat, although further studies are needed to explore this hypothesis. Loose iso- lation among populations could produce differences in song structure because birdsong is a cultur- ally transmitted trait. Key words: acoustic structure, Cettia diphone, geographic variation, Japanese bush warbler, Ogasawara Islands, song Parus major, within a habitat, suggesting that Introduction blue tits use species-specific songs (i.e., songs Like morphological traits, bird songs have with a trill) to reduce aggressive interactions evolved under various selection pressures from with great tits, whose songs never include trills ecological and social factors (Kroodsma and (Doutrelant and Lambrechts, 2001). -

Japan in Winter January 13–25, 2018
JAPAN IN WINTER JANUARY 13–25, 2018 Japanese (Red-crowned) Cranes dancing. Photo: S. Hilty LEADERS: KAZ SHINODA & STEVE HILTY with KOJI NIIYA one morning on HOKKAIDO LIST COMPILED BY: STEVE HILTY VICTOR EMANUEL NATURE TOURS, INC. 2525 WALLINGWOOD DRIVE, SUITE 1003 AUSTIN, TEXAS 78746 WWW.VENTBIRD.COM JAPAN IN WINTER: A CRANE & SEA-EAGLE SPECTACLE! By Steve Hilty One of the top highlights mentioned by most members of the group was a Ural Owl sleeping in a large, picturesque tree hollow. It was, in fact, an image that could have been plucked straight from an illustrated book of fairy tales from the Middle Ages. A male Eurasian Bullfinch in beautiful morning light also garnered top honors and, surprisingly, so did the diminutive Japanese Pygmy Woodpecker. For several of us, a large flock of Rooks eluding repeated prey- capture attempts by a Peregrine Falcon (the Rooks being more capable and wily than they might appear) over a large expanse of rice paddies was a trip highlight. Also prized were more than a dozen Stellar’s and White-tailed sea-eagles perched on a forested Hokkaido hillside during a snowstorm. The arrival of a Blakiston’s Fish-Owl at a small pool resulted in a mass exodus from our rather sedate and stylized Japanese dinner. And yes, then there were the Japanese Cranes, lumps of black and white fluff standing in a frigid river as steamy mists from the thermally- heated river water rose around them—a surreal and unforgettable setting. Surprisingly, perhaps, the Mandarin Ducks received not a single nod at the end—perhaps because they were a little distant—although they generated much excitement the morning we saw them, and the image of a stately pair cruising steadily across a mirror-smooth lake in early morning light, their narrow wake line trailing behind, will not likely be forgotten. -

Adobe PDF, Job 6
Noms français des oiseaux du Monde par la Commission internationale des noms français des oiseaux (CINFO) composée de Pierre DEVILLERS, Henri OUELLET, Édouard BENITO-ESPINAL, Roseline BEUDELS, Roger CRUON, Normand DAVID, Christian ÉRARD, Michel GOSSELIN, Gilles SEUTIN Éd. MultiMondes Inc., Sainte-Foy, Québec & Éd. Chabaud, Bayonne, France, 1993, 1re éd. ISBN 2-87749035-1 & avec le concours de Stéphane POPINET pour les noms anglais, d'après Distribution and Taxonomy of Birds of the World par C. G. SIBLEY & B. L. MONROE Yale University Press, New Haven and London, 1990 ISBN 2-87749035-1 Source : http://perso.club-internet.fr/alfosse/cinfo.htm Nouvelle adresse : http://listoiseauxmonde.multimania. -

Forest Bird Fauna of South China: Notes on Current Distribution and Status
FORKTAIL 22 (2006): 23–38 Forest bird fauna of South China: notes on current distribution and status LEE KWOK SHING, MICHAEL WAI-NENG LAU, JOHN R. FELLOWES and CHAN BOSCO PUI LOK From 1997 to 2004, a team from Hong Kong and southern China conducted rapid biodiversity surveys in 54 forest areas in the provinces of Guangdong, Guangxi and Hainan. A total of 372 bird species were recorded (201 in Guangdong, 299 in Guangxi and 164 in Hainan), including 12 globally threatened species, 50 China Key Protected Species and 44 species outside their previously recorded ranges. Breeding was confirmed for 94 species. In total, 232 species (62%) were recorded at five sites or fewer (2–10%). These include species at the edge of their range, migratory and wintering species inadequately sampled by these surveys, species more characteristic of non- forest habitats, and less conspicuous species that were under-recorded, but also rare and localised species. Of particular conservation concern are the globally threatened White-eared Night Heron Gorsachius magnificus, Cabot’s Tragopan Tragopan caboti, Hainan Partridge Arborophila ardens, White-necklaced Partridge Arborophila gingica, Fairy Pitta Pitta nympha, Pale-capped Pigeon Columba punicea, Brown-chested Jungle Flycatcher Rhinomyias brunneata and Gold-fronted Fulvetta Alcippe variegaticeps, and other species highly dependent on the region’s forests, such as Hainan Peacock Pheasant Polyplectron katsumatae, Pale-headed Woodpecker Gecinulus grantia, Blue-rumped Pitta Pitta soror, Swinhoe’s Minivet Pericrocotus cantonensis and Fujian Niltava Niltava davidi. At most of the sites visited, the main threat is habitat loss and degradation, especially clearance of natural forest for timber and agriculture; most remaining natural forests are fragmented and small in size. -

The Relationships and Evolution of the Southwest Pacific Warbler Genera I/I754 and Psama 7Hia’ (Sylviinae)
lIi’ ilson’ Bull., 95(2), 1983, pp. 184-198 THE RELATIONSHIPS AND EVOLUTION OF THE SOUTHWEST PACIFIC WARBLER GENERA I/I754 AND PSAMA 7HIA’ (SYLVIINAE) RONALD I. ORENSTEIN AND H. DOUGLAS PRATT The genera Vitia and Psamathia include three species of sylviine war- blers of forest undergrowth from Micronesia and Melanesia: the Fiji War- bler (V. rujkapilla) with races on Viti Levu (V. r. badiceps), Vanua Levu (V. r. castaneoptera), Taveuni (V. r. funebris), and Kadavu (V. r. rufica- pilla); the Shade Warbler (V. parens), a little-known bird of the mountains of San Cristobal, and probably Bougainville (Hadden 1981), in the Solomon Islands; and the Palau Bush-Warbler (Psamathia annae), restricted to Palau (Mayr 1945). Baker (1951:221) suggested, on the basis of external morphology, that P. annae was allied to Cettia diphone seebohmi of Luzon. Pratt et al. (1980) included the Palau bird in Cettia, and E. Mayr (in litt.) agrees with this treatment. The position of Vitia has been more obscure, although Delacour (194243) suggested that it, too, was allied to Cettia. Ramsay (1875), in his description of Vitia, compared V. ruJicapilla to Malurus (Malurinae) and Sericornis (Acanthizinae). Finsch (1876) placed V. r. (“Drymocha,era”) badiceps between Camaroptera and Orthotomus (Sylviinae). Mayr (1936) compared V. parens to Sericornis nouhuysi, but concluded that the two were not related. Morony et al. (1975) list Psa- mathia after Cettia, but place Vitia near the end of the Sylviinae with other genera of obscure affinity such as Ortygocichla and Megalurulus. This paper presents evidence that Vitia and Psamathia are Pacific island representatives of Cettia, closely related to C. -

Endemics & Winter Birds
Naturetrek Taiwan - Endemics & Winter Birds 14th November – 23rd November 2014 Gadwall Anas strepera Eurasian Wigeon Anas penelope American Wigeon Anas americana Eastern Spot-billed Duck Anas zonorhyncha Northern Shoveler Anas clypeata Northern Pintail Anas acuta Garganey Anas querquedula Baikal Teal Anas formosa Green-winged Teal Anas crecca Tufted Duck Aythya fuligula Chinese Bamboo-Partridge Bambusicola thoracicus Swinhoe's Pheasant Lophura swinhoii Mikado Pheasant Syrmaticus mikado Little Grebe Tachybaptus ruficollis Horned Grebe Podiceps auritus Great Cormorant Phalacrocorax carbo Yellow Bittern Ixobrychus sinensis Cinnamon Bittern Ixobrychus cinnamomeus Gray Heron Ardea cinerea Purple Heron Ardea purpurea Great Egret Ardea alba Intermediate Egret Mesophoyx intermedia Chinese Egret Egretta eulophotes Little Egret Egretta garzetta Pacific Reef-Heron Egretta sacra Pied Heron Egretta picata Cattle Egret Bubulcus ibis Chinese Pond-Heron Ardeola bacchus Striated Heron Butorides striata Black-crowned Night-Heron Nycticorax nycticorax Malayan Night-Heron Gorsachius melanolophus Sacred Ibis Threskiornis aethiopicus Black-headed Ibis Threskiornis melanocephalus Eurasian Spoonbill Platalea leucorodia Black-faced Spoonbill Platalea minor Osprey Pandion haliaetus Black-shouldered Kite Elanus caeruleus Oriental Honey-buzzard Pernis ptilorhynchus Crested Serpent-Eagle Spilornis cheela Mountain Hawk-Eagle Nisaetus nipalensis Black Eagle Ictinaetus malayensis Eastern Marsh-Harrier Circus spilonotus Crested Goshawk Accipiter trivirgatus Besra Accipiter -

List of Birds of Vietnam
(List of birds of Vietnam) No. ENGLISH NAME SCIENTIFIC NAME Note 1 Lesser Whistling-Duck Dendrocygna javanica 2 Bar-headed Goose Anser indicus Rare/Accidental 3 Graylag Goose Anser anser 4 Comb Duck Sarkidiornis melanotos 5 Ruddy Shelduck Tadorna ferruginea 6 Common Shelduck Tadorna tadorna 7 Cotton Pygmy-Goose Nettapus coromandelianus 8 Mandarin Duck Aix galericulata Rare/Accidental 9 Garganey Spatula querquedula 10 Northern Shoveler Spatula clypeata 11 Gadwall Mareca strepera 12 Falcated Duck Mareca falcata Near-threatened 13 Eurasian Wigeon Mareca penelope 14 Indian Spot-billed Duck Anas poecilorhyncha 15 Eastern Spot-billed Duck Anas zonorhyncha 16 Mallard Anas platyrhynchos Rare/Accidental 17 Northern Pintail Anas acuta 18 Green-winged Teal Anas crecca 19 White-winged Duck Asarcornis scutulata Endangered 20 Common Pochard Aythya ferina Vulnerable 21 Ferruginous Duck Aythya nyroca Near-threatened 22 Baer's Pochard Aythya baeri Critically endangered 23 Tufted Duck Aythya fuligula 24 Greater Scaup Aythya marila 25 Red-breasted Merganser Mergus serrator Rare/Accidental 26 Scaly-sided Merganser Mergus squamatus Endangered 27 Annam Partridge Arborophila merlini Endemic 28 Hill Partridge Arborophila torqueola 29 Rufous-throated Partridge Arborophila rufogularis 30 Bar-backed Partridge Arborophila brunneopectus 31 Orange-necked Partridge Arborophila davidi Endemic Near-threatened 32 Green-legged Patridge Arborophila chloropus 33 Crested Argus Rheinardia ocellata Near-threatened 34 Green Peafowl Pavo muticus Endangered 35 Germain's Peacock-Pheasant -
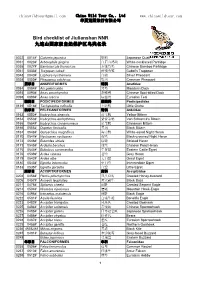
Bird Checklist of Jiulianshan NNR 九连山国家级自然保护区鸟类名录
[email protected] China Wild Tour Co,. Ltd www.chinawildtour.com 华夏荒野旅行有限公司 Bird checklist of Jiulianshan NNR 九连山国家级自然保护区鸟类名录 0022 0014# Coturnix japonica 鹌鹑 Japanese Quail 0031 0023# Arborophila gingica 白眉山鹧鸪 White-necklaced Partridge 0035 0027# Bambusicola thoracicus 灰胸竹鸡 Chinese Bamboo Partridge 0041 0033# Tragopan caboti 黄腹角雉 Cabot's Tragopan 0048 0040# Lophura nycthemera 白鹇 Silver Pheasant 0058 0050# Phasianus colchicus 雉鸡 Common Pheasant 雁形目 ANSERIFORMES 鸭科 Anatidae 0084 0084# Aix galericulata 鸳鸯 Mandarin Duck 0093 0090# Anas zonorhyncha 斑嘴鸭 Chinese Spot-billed Duck 0098 0096# Anas crecca 绿翅鸭 Eurasian Teal 䴙䴘目 PODICIPEDIFORMES 䴙䴘科 Podicipedidae 0139 0518# Tachybaptus ruficollis 小䴙䴘 Little Grebe 鹈形目 PELECANIFORMES 鹭科 Ardeidae 0163 0552# Ixobrychus sinensis 黄苇鳽 Yellow Bittern 0164 0553# Ixobrychus eurhythmus 紫背苇鳽 Von Schrenck's Bittern 0165 0554# Ixobrychus cinnamomeus 栗苇鳽 Cinnamon Bittern 0166 0555# Dupetor flavicollis 黑鳽 Black Bittern 0167 0548# Gorsachius magnificus 海南鳽 White-eared Night Heron 0170 0547# Nycticorax nycticorax 夜鹭 Black-crowned Night Heron 0172 0546# Butorides striata 绿鹭 Striated Heron 0173 0545# Ardeola bacchus 池鹭 Chinese Pond Heron 0175 0544# Bubulcus coromandus 牛背鹭 Eastern Cattle Egret 0176 0539# Ardea cinerea 苍鹭 Grey Heron 0179 0542# Ardea alba 大白鹭 Great Egret 0180 0543# Egretta intermedia 中白鹭 Intermediate Egret 0183 0535# Egretta garzetta 白鹭 Little Egret 鹰形目 ACCIPITRIFORMES 鹰科 Accipitridae 0204 0458# Pernis ptilorhynchus 凤头蜂鹰 Crested Honey-buzzard 0206 0457# Aviceda leuphotes 黑冠鹃隼 Black Baza 0211 0475# -

Mitochondrial Phylogeny of Locustella and Related Genera
JOURNAL OF AVIAN BIOLOGY 35: 105Á/110, 2004 Mitochondrial phylogeny of Locustella and related genera Sergei V. Drovetski, Robert M. Zink, Igor V. Fadeev, Evgeniy V. Nesterov, Evgeniy A. Koblik, Yaroslav A. Red’kin and Sievert Rohwer Drovetski, S. V., Zink, R. M., Fadeev I. V., Nesterov, E. V., Koblik E. A., Red’kin, Y. A. and Rohwer, S. 2004. Mitochondrial phylogeny of Locustella and related genera. */ J. Avian Biol. 35: 105Á/110. We used maximum likelihood analysis of complete mitochondrial ND2 sequences (1041 bp) to clarify the taxonomy and relationships of various species and genera of grass and bush warblers. The tree revealed two clades of grass and bush warblers. One clade was comprised of all four western Palearctic Locustella and two species of Asian Bradypterus. The other clade included five eastern Palearctic Locustella (including the distinctive Sakhalin warbler Locustella amnicola) and the marsh grassbird Megalurus pryeri. African Bradypterus and Australian little grassbird Megalurus gramineus were distantly related to their Asian congeners. Therefore, current taxonomy of these genera does not reflect their evolutionary history and needs revision. It is proposed that a phylogenetic analysis of morphology and ecological preferences would show that the current taxonomy of grass and bush warblers reflects species’ habitat preferences and morphology related to locomotion and foraging in their habitats, rather than their shared ancestry. Distinct clades were found in grasshopper warbler Locustella naevia and Pallas’s grasshopper warbler L. certhiola. Detailed phylogeographic studies are needed to elucidate the species status of the clades within these two species. S. V. Drovetski (correspondence), Department of Ecology, Evolution and Behavior, University of Minnesota, 100 Ecology Building, 1987 Upper Buford Circle, St.