Full Text in Pdf Format
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Biostratigraphic Precision of the Cruziana Rugosa Group: a Study from the Ordovician Succession of Southern and Central Bolivia
Geol. Mag. 144 (2), 2007, pp. 289–303. c 2007 Cambridge University Press 289 doi:10.1017/S0016756807003093 First published online 9 February 2007 Printed in the United Kingdom Biostratigraphic precision of the Cruziana rugosa group: a study from the Ordovician succession of southern and central Bolivia SVEN O. EGENHOFF∗, BERND WEBER†, OLIVER LEHNERT‡ &JORG¨ MALETZ§ ∗Colorado State University, Department of Geosciences, 322 Natural Resources Building, Fort Collins, CO 80523-1482, USA †Freie Universitat¨ Berlin, Institut fur¨ Geologische Wissenschaften, Fachrichtung Geologie, Malteserstrasse 74-100, D-12249 Berlin, Germany ‡University of Erlangen, Institute of Geology and Mineralogie, Schlossgarten 5, D-91054 Erlangen, Germany §Department of Geology, State University of New York at Buffalo, 772 Natural Sciences and Mathematics Complex, Buffalo, New York 14260-3050, USA (Received 10 October 2005; revised version received 1 May 2006; accepted 22 May 2006) Abstract – Cruziana ichnospecies have been repeatedly reported to have biostratigraphic significance. This study presents a re-evaluation of the arthropod ichnotaxa of the Cruziana rugosa Group from bio- and/or lithostratigraphically well-defined Lower to Upper Ordovician siliciclastic sections of southern and central Bolivia. With the exception of Cruziana rouaulti, the ichnofaunas contain all the members of the Cruziana rugosa Group throughout the Ordovician (Arenig to Caradoc) successions in Bolivia. The Bolivian material therefore indicates that these arthropod ichnofossil assemblages are suitable for recognizing Ordovician strata in Bolivia. These findings cast doubt on their use as reliable indicators for a global intra-Ordovician (Arenig to Caradoc) biozonation of Peri-Gondwanan sedimentary successions. Keywords: Cruziana, biostratigraphy, Bolivia, Ordovician. 1. Introduction to the present study. -

Königreichs Zur Abgrenzung Der Der Kommission in Übereinstimmung
19 . 5 . 75 Amtsblatt der Europäischen Gemeinschaften Nr . L 128/23 1 RICHTLINIE DES RATES vom 28 . April 1975 betreffend das Gemeinschaftsverzeichnis der benachteiligten landwirtschaftlichen Gebiete im Sinne der Richtlinie 75/268/EWG (Vereinigtes Königreich ) (75/276/EWG ) DER RAT DER EUROPAISCHEN 1973 nach Abzug der direkten Beihilfen, der hill GEMEINSCHAFTEN — production grants). gestützt auf den Vertrag zur Gründung der Euro Als Merkmal für die in Artikel 3 Absatz 4 Buch päischen Wirtschaftsgemeinschaft, stabe c ) der Richtlinie 75/268/EWG genannte ge ringe Bevölkerungsdichte wird eine Bevölkerungs gestützt auf die Richtlinie 75/268/EWG des Rates ziffer von höchstens 36 Einwohnern je km2 zugrunde vom 28 . April 1975 über die Landwirtschaft in Berg gelegt ( nationaler Mittelwert 228 , Mittelwert in der gebieten und in bestimmten benachteiligten Gebie Gemeinschaft 168 Einwohner je km2 ). Der Mindest ten (*), insbesondere auf Artikel 2 Absatz 2, anteil der landwirtschaftlichen Erwerbspersonen an der gesamten Erwerbsbevölkerung beträgt 19 % auf Vorschlag der Kommission, ( nationaler Mittelwert 3,08 % , Mittelwert in der Gemeinschaft 9,58 % ). nach Stellungnahme des Europäischen Parlaments , Eigenart und Niveau der vorstehend genannten nach Stellungnahme des Wirtschafts- und Sozialaus Merkmale, die von der Regierung des Vereinigten schusses (2 ), Königreichs zur Abgrenzung der der Kommission mitgeteilten Gebiete herangezogen wurden, ent sprechen den Merkmalen der in Artikel 3 Absatz 4 in Erwägung nachstehender Gründe : der Richtlinie -

001-012 Primeras Páginas
PUBLICACIONES DEL INSTITUTO GEOLÓGICO Y MINERO DE ESPAÑA Serie: CUADERNOS DEL MUSEO GEOMINERO. Nº 9 ADVANCES IN TRILOBITE RESEARCH ADVANCES IN TRILOBITE RESEARCH IN ADVANCES ADVANCES IN TRILOBITE RESEARCH IN ADVANCES planeta tierra Editors: I. Rábano, R. Gozalo and Ciencias de la Tierra para la Sociedad D. García-Bellido 9 788478 407590 MINISTERIO MINISTERIO DE CIENCIA DE CIENCIA E INNOVACIÓN E INNOVACIÓN ADVANCES IN TRILOBITE RESEARCH Editors: I. Rábano, R. Gozalo and D. García-Bellido Instituto Geológico y Minero de España Madrid, 2008 Serie: CUADERNOS DEL MUSEO GEOMINERO, Nº 9 INTERNATIONAL TRILOBITE CONFERENCE (4. 2008. Toledo) Advances in trilobite research: Fourth International Trilobite Conference, Toledo, June,16-24, 2008 / I. Rábano, R. Gozalo and D. García-Bellido, eds.- Madrid: Instituto Geológico y Minero de España, 2008. 448 pgs; ils; 24 cm .- (Cuadernos del Museo Geominero; 9) ISBN 978-84-7840-759-0 1. Fauna trilobites. 2. Congreso. I. Instituto Geológico y Minero de España, ed. II. Rábano,I., ed. III Gozalo, R., ed. IV. García-Bellido, D., ed. 562 All rights reserved. No part of this publication may be reproduced or transmitted in any form or by any means, electronic or mechanical, including photocopy, recording, or any information storage and retrieval system now known or to be invented, without permission in writing from the publisher. References to this volume: It is suggested that either of the following alternatives should be used for future bibliographic references to the whole or part of this volume: Rábano, I., Gozalo, R. and García-Bellido, D. (eds.) 2008. Advances in trilobite research. Cuadernos del Museo Geominero, 9. -

Introduction to the Trilobites: Morphology, Ecology, Macroevolution and More by Michelle M
Introduction to the Trilobites: Morphology, Ecology, Macroevolution and More By Michelle M. Casey1, Perry Kennard2, and Bruce S. Lieberman1, 3 1Biodiversity Institute, University of Kansas, Lawrence, KS, 66045, 2Earth Science Teacher, Southwest Middle School, USD497, and 3Department of Ecology and Evolutionary Biology, University of Kansas, Lawrence, KS 66045 Middle level laboratory exercise for Earth or General Science; supported provided by National Science Foundation (NSF) grants DEB-1256993 and EF-1206757. Learning Goals and Pedagogy This lab is designed for middle level General Science or Earth Science classes. The learning goals for this lab are the following: 1) to familiarize students with the anatomy and terminology relating to trilobites; 2) to give students experience identifying morphologic structures on real fossil specimens 3) to highlight major events or trends in the evolutionary history and ecology of the Trilobita; and 4) to expose students to the study of macroevolution in the fossil record using trilobites as a case study. Introduction to the Trilobites The Trilobites are an extinct subphylum of the Arthropoda (the most diverse phylum on earth with nearly a million species described). Arthropoda also contains all fossil and living crustaceans, spiders, and insects as well as several other extinct groups. The trilobites were an extremely important and diverse type of marine invertebrates that lived during the Paleozoic Era. They only lived in the oceans but occurred in all types of marine environments, and ranged in size from less than a centimeter to almost a meter across. They were once one of the most successful of all animal groups and in certain fossil deposits, especially in the Cambrian, Ordovician, and Devonian periods, they are extremely abundant. -

Zapla Range, Subandean Ranges, Jujuy Province
Zapla Range, Subandean Ranges, Jujuy Province Gladys Ortega1, M. Cristina Moya2 and Guillermo L. Albanesi1 1 CONICET. Museo de Paleontología, Universidad Nacional de Córdoba, C.C. 1598, 5000 Córdoba. E–mail: [email protected] / [email protected] 2 CONICET, CIUNSA. Universidad Nacional de Salta. Buenos Aires 177, 4400 Salta. e–mail:[email protected] Introduction Stratigraphic framework The Subandean Ranges (Sierras Subandinas) are placed between the Eastern Cordillera (Cordillera Oriental) to the west and Chaco–pampean Plain to the east, in northwestern Argentina. Stratigraphic and structural studies of this geological province were carried out by Bonarelli (1913, 1921), Baldis et al.(1975), Mingramm et al. (1979), and Ramos (1999), among others. Ancient deposits of the Subandean Ranges correspond to the Proterozoic and Ordovician System, which are restricted to the western part. Silurian–Devonian rocks of wider distribution integrate a thick marine–deltaic succession tapering to the east, which is linked to a foreland basin (Turner, 1967; Ramos, 1999). The Ocloyic unconformity (Turner & Méndez, 1975; Ramos, 1986) separates Lower–Middle Ordovician rocks from Hirnantian and younger deposits (Moya, 1999). The Lower Paleozoic succession is covered by thick marine and continental sequences of Neopaleozoic, Mesozoic and Cenozoic ages. An angular unconformity is present between ancient deposits and Miocene to Quaternary sediments. The Subandean Ranges show wide east–vergence anticlines, limited by thrusts and overthrusts, whose detachment levels are Silurian–Devonian shales (Ramos, 1999). The structural style of this geological province allows for the identification of the Interandean System to the west, and the Subandean System sensu stricto to the east (Ramos, op. -

Cruziana Traces from the Late Silurian (Pridoli) Carbonate Shelf of Saaremaa, Estonia
Estonian Journal of Earth Sciences, 2014, 63, 2, 71–75 doi: 10.3176/earth.2014.06 Cruziana traces from the Late Silurian (Pridoli) carbonate shelf of Saaremaa, Estonia Olev Vinn Department of Geology, University of Tartu, Ravila 14A, 50411 Tartu, Estonia; [email protected] Received 25 February 2014, accepted 28 April 2014 Abstract. Late Pridoli Cruziana traces have recently been found in carbonate shelf sediments of the Ohesaare Formation on Saaremaa Island, Estonia. Cruziana isp. is interpreted here as a locomotory trace (repichnia) of an arthropod, possibly a trilobite. Cruziana traces previously known from the Silurian of Baltica differ from Cruziana isp., indicating that the diversity of Cruziana traces in the late Silurian of Baltica was higher than previously thought. Key words: trace fossils, Late Silurian, carbonate platform, Cruziana, trilobites, Baltica. INTRODUCTION Cruziana traces have mostly been restricted to specific palaeogeographic regions and localities (Seilacher 2007). Trace fossils of the Silurian of Estonia and the eastern Cruziana kufraensis has been described from Lower Baltic in general are poorly studied. A few reports are Silurian rocks of Libya and C. acacensis from the Lower devoted to borings in the hard substrates of the Silurian Silurian of North Africa and South America (Seilacher of Estonia (Vinn & Wilson 2010a, 2010b). Even less 2007). Cruziana pudica has a wide distribution in the is known about the softground substrate trace fossils. Upper Ordovician and Lower Silurian of North America Only Skolithos burrows have previously been described (Garlock & Isaacson 1977; Seilacher 2007). Records of from the late Pridoli of Saaremaa (Vinn & Wilson Rhusophycus pudicum and Rhusophycus pudica should 2013). -

Death in the Shallows the Record of Permo-Triassic Mass Extinction In
University of Birmingham Death in the shallows Wignall, Paul; Chu, Daoliang; Hilton, Jason; Dal Corso, Jacopo; Wu, Yuyang; Wang, Yao; Atkinson, Jed; Tong, Jinnan DOI: 10.1016/j.gloplacha.2020.103176 License: Creative Commons: Attribution-NonCommercial-NoDerivs (CC BY-NC-ND) Document Version Publisher's PDF, also known as Version of record Citation for published version (Harvard): Wignall, P, Chu, D, Hilton, J, Dal Corso, J, Wu, Y, Wang, Y, Atkinson, J & Tong, J 2020, 'Death in the shallows: the record of Permo-Triassic mass extinction in paralic settings, southwest China', Global and Planetary Change, vol. 189, 103176, pp. 1-12. https://doi.org/10.1016/j.gloplacha.2020.103176 Link to publication on Research at Birmingham portal General rights Unless a licence is specified above, all rights (including copyright and moral rights) in this document are retained by the authors and/or the copyright holders. The express permission of the copyright holder must be obtained for any use of this material other than for purposes permitted by law. •Users may freely distribute the URL that is used to identify this publication. •Users may download and/or print one copy of the publication from the University of Birmingham research portal for the purpose of private study or non-commercial research. •User may use extracts from the document in line with the concept of ‘fair dealing’ under the Copyright, Designs and Patents Act 1988 (?) •Users may not further distribute the material nor use it for the purposes of commercial gain. Where a licence is displayed above, please note the terms and conditions of the licence govern your use of this document. -

Th TRILO the Back to the Past Museum Guide to TRILO BITES
With regard to human interest in fossils, trilobites may rank second only to dinosaurs. Having studied trilobites most of my life, the English version of The Back to the Past Museum Guide to TRILOBITES by Enrico Bonino and Carlo Kier is a pleasant treat. I am captivated by the abundant color images of more than 600 diverse species of trilobites, mostly from the authors’ own collections. Carlo Kier The Back to the Past Museum Guide to Specimens amply represent famous trilobite localities around the world and typify forms from most of the Enrico Bonino Enrico 250-million-year history of trilobites. Numerous specimens are masterpieces of modern professional preparation. Richard A. Robison Professor Emeritus University of Kansas TRILOBITES Enrico Bonino was born in the Province of Bergamo in 1966 and received his degree in Geology from the Depart- ment of Earth Sciences at the University of Genoa. He currently lives in Belgium where he works as a cartographer specialized in the use of satellite imaging and geographic information systems (GIS). His proficiency in the use of digital-image processing, a healthy dose of artistic talent, and a good knowledge of desktop publishing software have provided him with the skills he needed to create graphics, including dozens of posters and illustrations, for all of the displays at the Back to the Past Museum in Cancún. In addition to his passion for trilobites, Enrico is particularly inter- TRILOBITES ested in the life forms that developed during the Precambrian. Carlo Kier was born in Milan in 1961. He holds a degree in law and is currently the director of the Azul Hotel chain. -

An Inventory of Trilobites from National Park Service Areas
Sullivan, R.M. and Lucas, S.G., eds., 2016, Fossil Record 5. New Mexico Museum of Natural History and Science Bulletin 74. 179 AN INVENTORY OF TRILOBITES FROM NATIONAL PARK SERVICE AREAS MEGAN R. NORR¹, VINCENT L. SANTUCCI1 and JUSTIN S. TWEET2 1National Park Service. 1201 Eye Street NW, Washington, D.C. 20005; -email: [email protected]; 2Tweet Paleo-Consulting. 9149 79th St. S. Cottage Grove. MN 55016; Abstract—Trilobites represent an extinct group of Paleozoic marine invertebrate fossils that have great scientific interest and public appeal. Trilobites exhibit wide taxonomic diversity and are contained within nine orders of the Class Trilobita. A wealth of scientific literature exists regarding trilobites, their morphology, biostratigraphy, indicators of paleoenvironments, behavior, and other research themes. An inventory of National Park Service areas reveals that fossilized remains of trilobites are documented from within at least 33 NPS units, including Death Valley National Park, Grand Canyon National Park, Yellowstone National Park, and Yukon-Charley Rivers National Preserve. More than 120 trilobite hototype specimens are known from National Park Service areas. INTRODUCTION Of the 262 National Park Service areas identified with paleontological resources, 33 of those units have documented trilobite fossils (Fig. 1). More than 120 holotype specimens of trilobites have been found within National Park Service (NPS) units. Once thriving during the Paleozoic Era (between ~520 and 250 million years ago) and becoming extinct at the end of the Permian Period, trilobites were prone to fossilization due to their hard exoskeletons and the sedimentary marine environments they inhabited. While parks such as Death Valley National Park and Yukon-Charley Rivers National Preserve have reported a great abundance of fossilized trilobites, many other national parks also contain a diverse trilobite fauna. -

The Silurian of Central Kentucky, U.S.A.: Stratigraphy, Palaeoenvironments and Palaeoecology
The Silurian of central Kentucky, U.S.A.: Stratigraphy, palaeoenvironments and palaeoecology FRANK R. ETTENSOHN, R. THOMAS LIERMAN, CHARLES E. MASON, WILLIAM M. ANDREWS, R. TODD HENDRICKS, DANIEL J. PHELPS & LAWRENCE A. GORDON ETTENSOHN, F.R., LIERMAN, R.T., MASON, C.E., ANDREWS, W.M., HENDRICKS, R.T., PHELPS, D.J. & GORDON, L.A., 2013:04:26. The Silurian of central Kentucky, U.S.A.: Stratigraphy, palaeoenvironments and palaeoecology. Memoirs of the Association of Australasian Palaeontologists 44, 159-189. ISSN 0810-8889. Silurian rocks in Kentucky are exposed on the eastern and western flanks of the Cincinnati Arch, a large-wavelength cratonic structure separating the Appalachian foreland basin from the intracratonic Illinois Basin. The Cincinnati Arch area experienced uplift during latest Ordovician-early Silurian time, so that the exposed Silurian section is relatively thin due to onlap and post- Silurian erosional truncation on the arch. On both flanks of the arch, dolomitic carbonates predominate, but the section on the eastern side reflects a more shale-rich ramp that faced eastern Appalachian source areas. In the Silurian section on the western side of the arch, which apparently developed across a platform-like isolation-accommodation zone, shales are rare except dur- ing some highstand episodes, and rocks in the area reflect deposition across a broad, low-gradient shelf area, interrupted by structurally controlled topographic breaks. Using the progression of interpreted depositional environments and nearshore faunal communities, a relative sea-level curve, which parallels those of previous workers, was generated for the section in Kentucky. While the curve clearly shows the influence of glacial eustasy, distinct indications of the far-field, flexural influence of Taconian and Salinic tectonism are also present. -
2012 Gwynedd
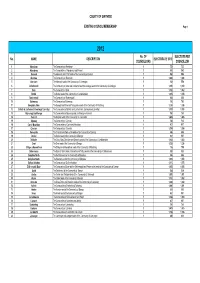
COUNTY OF GWYNEDD EXISTING COUNCIL MEMBERSHIP Page 1 2012 No. OF ELECTORS PER No. NAME DESCRIPTION ELECTORATE 2012 COUNCILLORS COUNCILLOR 1 Aberdaron The Community of Aberdaron 1 733 733 2 Aberdovey The Communities of Aberdovey and Pennal 1 960 960 3 Abererch The Abererch and Y Ffôr wards of the Community of Llannor 1 998 998 4 Abermaw The Community of Barmouth 1 1,608 1,608 5 Abersoch The Abersoch ward of the Community of Llanengan 1 558 558 6 Arllechwedd The Community of Aber and Llanllechid and the Llandygai ward of the Community of Llandygai 1 1,010 1,010 7 Bala The Community of Bala 1 1,362 1,362 8 Bethel The Bethel ward of the Community of Llanddeiniolen 1 1,015 1,015 9 Bontnewydd The Community of Bontnewydd 1 836 836 10 Botwnnog The Community of Botwnnog 1 700 700 11 Bowydd & Rhiw The Bowydd and Rhiw and Tanygrisiau wards of the Community of Ffestiniog 1 1,218 1,218 12 Brithdir & Llanfachreth/ Ganllwyd/ Llanelltyd The Communities of Brithdir and Llanfachreth, Ganllwyd and Llanelltyd 1 1,103 1,103 13 Bryn-crug/ Llanfihangel The Communities of Bryn-crug and Llanfihangel-y-Pennant 1 761 761 14 Cadnant The Dwyrain ward of the Community of Caernarfon 1 1,405 1,405 15 Clynnog The Community of Clynnog 1 723 723 16 Corris/ Mawddwy The Communities of Corris and Mawddwy 1 917 917 17 Criccieth The Community of Criccieth 1 1,354 1,354 18 Cwm-y-Glo The Ceunant and Cwm-y-Glo wards of the Community of Llanrug 1 696 696 19 Deiniol The Deiniol ward of the Community of Bangor 1 537 537 20 Deiniolen The Clwt y Bont, Deiniolen and Dinorwic wards of -

The Parliamentary Constituencies and Assembly Electoral Regions (Wales) Order 2006
STATUTORY INSTRUMENTS 2006 No. 1041 REPRESENTAION OF THE PEOPLE, WALES REDISTRIBUTION OF SEATS The Parliamentary Constituencies and Assembly Electoral Regions (Wales) Order 2006 Made - - - - - 11th April 2006 Coming into force in accordance with article 1(2) £5.50 STATUTORY INSTRUMENTS 2006 No. 1041 REPRESENTATION OF THE PEOPLE, WALES REDISTRIBUTION OF SEATS The Parliamentary Constituencies and Assembly Electoral Regions (Wales) Order 2006 Made - - - - 11th April 2006 Coming into force in accordance with article 1(2) At the Court at Windsor Castle, the 11th day of April Present, The Queen’s Most Excellent Majesty in Council The Boundary Commission for Wales (“the Commission”) have, in accordance with section 3(1) of the Parliamentary Constituencies Act 1986(a), submitted to the Secretary of State a report dated 31st January 2005(b) showing the parliamentary constituencies into which they recommend, in accordance with that Act, that Wales should be divided. That report also shows, as provided for by paragraph 7(2) of Schedule 1 to the Government of Wales Act 1998(c), the alterations in the electoral regions of the National Assembly for Wales which the Commission recommend(d). A draft Order in Council together with a copy of the Commission’s report was laid before Parliament by the Secretary of State to give effect, without modifications, to the recommendations contained in the report, and each House of Parliament has by resolution approved that draft. Now, therefore, Her Majesty, is pleased, by and with the advice of Her Privy Council, to make the following Order under section 4 of the Parliamentary Constituencies Act 1986: (a) 1986 c.56.