Download Poster
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

The Role of Movements in the Development of Sutural
J. Anat. (1983), 137, 3, pp. 591-599 591 With 11 figures Printed in Great Britain The role of movements in the development of sutural and diarthrodial joints tested by long-term paralysis of chick embryos MAURITS PERSSON Department of Orthodontics, Faculty of Odontology, University of Umea, Norrlandsgatan 18 B, S-902 48 Umea, Sweden (Accepted 17 February 1983) INTRODUCTION The factors regulating the development of sutural fibrous joints are a subject of controversy. Movements of muscular origin (Pritchard, Scott & Girgis, 1956), an osteogenesis-inhibiting factor (Markens, 1975) and physiolQgical cell death (Ten Cate, Freeman & Dickinson, 1977) have been suggested. More recently, Smith & Tondury (1978) concluded that stretch-growth tensile forces in the dura mater are responsible for the development of the calvaria and its sutures, the primary deter- minant in their development being the form and growth of the early brain. A similar biomechanical explanation for the morphogenesis of sutures in areas of the skull where no dura mater exists was also given by Persson & Roy (1979). Based on studies of suture development and bony fusion in the rabbit palate, they con- cluded that the spatial separation of bones during growth is the factor regulating suture formation. Lack of such movements of developing bones results in bone fusion when the bones meet. Further, when cranial bones at suture sites are im- mobilised, fusion of the bones across the suture is produced (Persson et al. 1979). In the normal development of diarthrodial joints, skeletal muscle contractions are essential (Murray & Selby, 1930; Lelkes, 1958; Drachman & Coulombre, 1962; Murray & Drachman, 1969; Hall, 1975), and lack of muscular movements results in stiff, fused joints. -

Hands-On Human Evolution: a Laboratory Based Approach
Hands-on Human Evolution: A Laboratory Based Approach Developed by Margarita Hernandez Center for Precollegiate Education and Training Author: Margarita Hernandez Curriculum Team: Julie Bokor, Sven Engling A huge thank you to….. Contents: 4. Author’s note 5. Introduction 6. Tips about the curriculum 8. Lesson Summaries 9. Lesson Sequencing Guide 10. Vocabulary 11. Next Generation Sunshine State Standards- Science 12. Background information 13. Lessons 122. Resources 123. Content Assessment 129. Content Area Expert Evaluation 131. Teacher Feedback Form 134. Student Feedback Form Lesson 1: Hominid Evolution Lab 19. Lesson 1 . Student Lab Pages . Student Lab Key . Human Evolution Phylogeny . Lab Station Numbers . Skeletal Pictures Lesson 2: Chromosomal Comparison Lab 48. Lesson 2 . Student Activity Pages . Student Lab Key Lesson 3: Naledi Jigsaw 77. Lesson 3 Author’s note Introduction Page The validity and importance of the theory of biological evolution runs strong throughout the topic of biology. Evolution serves as a foundation to many biological concepts by tying together the different tenants of biology, like ecology, anatomy, genetics, zoology, and taxonomy. It is for this reason that evolution plays a prominent role in the state and national standards and deserves thorough coverage in a classroom. A prime example of evolution can be seen in our own ancestral history, and this unit provides students with an excellent opportunity to consider the multiple lines of evidence that support hominid evolution. By allowing students the chance to uncover the supporting evidence for evolution themselves, they discover the ways the theory of evolution is supported by multiple sources. It is our hope that the opportunity to handle our ancestors’ bone casts and examine real molecular data, in an inquiry based environment, will pique the interest of students, ultimately leading them to conclude that the evidence they have gathered thoroughly supports the theory of evolution. -

Is the Skeleton Male Or Female? the Pelvis Tells the Story
Activity: Is the Skeleton Male or Female? The pelvis tells the story. Distinct features adapted for childbearing distinguish adult females from males. Other bones and the skull also have features that can indicate sex, though less reliably. In young children, these sex-related features are less obvious and more difficult to interpret. Subtle sex differences are detectable in younger skeletons, but they become more defined following puberty and sexual maturation. What are the differences? Compare the two illustrations below in Figure 1. Female Pelvic Bones Male Pelvic Bones Broader sciatic notch Narrower sciatic notch Raised auricular surface Flat auricular surface Figure 1. Female and male pelvic bones. (Source: Smithsonian Institution, illustrated by Diana Marques) Figure 2. Pelvic bone of the skeleton in the cellar. (Source: Smithsonian Institution) Skull (Cranium and Mandible) Male Skulls Generally larger than female Larger projections behind the Larger brow ridges, with sloping, ears (mastoid processes) less rounded forehead Square chin with a more vertical Greater definition of muscle (acute) angle of the jaw attachment areas on the back of the head Figure 3. Male skulls. (Source: Smithsonian Institution, illustrated by Diana Marques) Female Skulls Smoother bone surfaces where Smaller projections behind the muscles attach ears (mastoid processes) Less pronounced brow ridges, Chin more pointed, with a larger, with more vertical forehead obtuse angle of the jaw Sharp upper margins of the eye orbits Figure 4. Female skulls. (Source: Smithsonian Institution, illustrated by Diana Marques) What Do You Think? Comparing the skull from the cellar in Figure 5 (below) with the illustrated male and female skulls in Figures 3 and 4, write Male or Female to note the sex depicted by each feature. -

A Description of the Geological Context, Discrete Traits, and Linear Morphometrics of the Middle Pleistocene Hominin from Dali, Shaanxi Province, China
AMERICAN JOURNAL OF PHYSICAL ANTHROPOLOGY 150:141–157 (2013) A Description of the Geological Context, Discrete Traits, and Linear Morphometrics of the Middle Pleistocene Hominin from Dali, Shaanxi Province, China Xinzhi Wu1 and Sheela Athreya2* 1Laboratory for Human Evolution, Institute of Vertebrate Paleontology and Paleoanthropology, Chinese Academy of Sciences, Beijing 100044, China 2Department of Anthropology, Texas A&M University, College Station, TX 77843 KEY WORDS Homo heidelbergensis; Homo erectus; Asia ABSTRACT In 1978, a nearly complete hominin Afro/European Middle Pleistocene Homo and align it fossil cranium was recovered from loess deposits at the with Asian H. erectus.Atthesametime,itdisplaysa site of Dali in Shaanxi Province, northwestern China. more derived morphology of the supraorbital torus and It was subsequently briefly described in both English supratoral sulcus and a thinner tympanic plate than and Chinese publications. Here we present a compre- H. erectus, a relatively long upper (lambda-inion) occi- hensive univariate and nonmetric description of the pital plane with a clear separation of inion and opis- specimen and provide comparisons with key Middle thocranion, and an absolute and relative increase in Pleistocene Homo erectus and non-erectus hominins brain size, all of which align it with African and Euro- from Eurasia and Africa. In both respects we find pean Middle Pleistocene Homo. Finally, traits such as affinities with Chinese H. erectus as well as African the form of the frontal keel and the relatively short, and European Middle Pleistocene hominins typically broad midface align Dali specifically with other referred to as Homo heidelbergensis.Specifically,the Chinese specimens from the Middle Pleistocene Dali specimen possesses a low cranial height, relatively and Late Pleistocene, including H. -

Cranial Suture Mesenchymal Stem Cells: Insights and Advances
biomolecules Review Cranial Suture Mesenchymal Stem Cells: Insights and Advances Bo Li 1, Yigan Wang 1, Yi Fan 2, Takehito Ouchi 3 , Zhihe Zhao 1,* and Longjiang Li 4,* 1 State Key Laboratory of Oral Diseases, National Clinical Research Center for Oral Diseases, Department of Orthodontics, West China Hospital of Stomatology, Sichuan University, Chengdu 610041, China; [email protected] (B.L.); [email protected] (Y.W.) 2 State Key Laboratory of Oral Diseases, National Clinical Research Center for Oral Diseases, Department of Cariology and Endodontics, West China Hospital of Stomatology, Sichuan University, Chengdu 610041, China; [email protected] 3 Department of Physiology, Tokyo Dental College, Tokyo 1010061, Japan; [email protected] 4 State Key Laboratory of Oral Diseases, National Clinical Research Center for Oral Diseases, Department of Head and Neck Oncology, West China Hospital of Stomatology, Sichuan University, Chengdu 610041, China * Correspondence: [email protected] (Z.Z.); [email protected] (L.L.) Abstract: The cranial bones constitute the protective structures of the skull, which surround and protect the brain. Due to the limited repair capacity, the reconstruction and regeneration of skull defects are considered as an unmet clinical need and challenge. Previously, it has been proposed that the periosteum and dura mater provide reparative progenitors for cranial bones homeostasis and injury repair. In addition, it has also been speculated that the cranial mesenchymal stem cells reside in the perivascular niche of the diploe, namely, the soft spongy cancellous bone between the interior and exterior layers of cortical bone of the skull, which resembles the skeletal stem cells’ distribution pattern of the long bone within the bone marrow. -

Ectocranial Suture Closure in Pan Troglodytes and Gorilla Gorilla: Pattern and Phylogeny James Cray Jr.,1* Richard S
AMERICAN JOURNAL OF PHYSICAL ANTHROPOLOGY 136:394–399 (2008) Ectocranial Suture Closure in Pan troglodytes and Gorilla gorilla: Pattern and Phylogeny James Cray Jr.,1* Richard S. Meindl,2 Chet C. Sherwood,3 and C. Owen Lovejoy2 1Department of Anthropology, University of Pittsburgh, Pittsburgh, PA 15260 2Department of Anthropology and Division of Biomedical Sciences, Kent State University, Kent, OH 44242 3Department of Anthropology, The George Washington University, Washington, DC 20052 KEY WORDS cranial suture; synostosis; variation; phylogeny; Guttman analysis ABSTRACT The order in which ectocranial sutures than either does with G. gorilla, we hypothesized that this undergo fusion displays species-specific variation among phylogenetic relationship would be reflected in the suture primates. However, the precise relationship between suture closure patterns of these three taxa. Results indicated that closure and phylogenetic affinities is poorly understood. In while all three species do share a similar lateral-anterior this study, we used Guttman Scaling to determine if the closure pattern, G. gorilla exhibits a unique vault pattern, modal progression of suture closure differs among Homo which, unlike humans and P. troglodyte s, follows a strong sapiens, Pan troglodytes,andGorilla gorilla.BecauseDNA posterior-to-anterior gradient. P. troglodytes is therefore sequence homologies strongly suggest that P. tr og lodytes more like Homo sapiens in suture synostosis. Am J Phys and Homo sapiens share a more recent common ancestor Anthropol 136:394–399, 2008. VC 2008 Wiley-Liss, Inc. The biological basis of suture synostosis is currently Morriss-Kay et al. (2001) found that maintenance of pro- poorly understood, but appears to be influenced by a liferating osteogenic stem cells at the margins of mem- combination of vascular, hormonal, genetic, mechanical, brane bones forming the coronal suture requires FGF and local factors (see review in Cohen, 1993). -
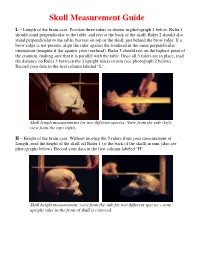
AP Skull Measurement Guide, Laminate
Skull Measurement Guide L – Length of the brain case: Position three rulers as shown in photograph 1 below. Ruler 1 should stand perpendicular to the table, and rest at the back of the skull. Ruler 2 should also stand perpendicular to the table, but rest on top of the skull, just behind the brow ridge. If a brow ridge is not present, align the ruler against the forehead in the same perpendicular orientation (imagine it flat against your forehead). Ruler 3 should rest on the highest point of the cranium, making sure that it is parallel with the table. Once all 3 rulers are in place, read the distance on Ruler 3 between the 2 upright rulers in mm (see photograph 2 below). Record your data in the first column labeled “L”. Skull length measurements for two different species: View from the side (left), view from the top (right). H – Height of the brain case: Without moving the 3 rulers from your measurement of Length, read the height of the skull off Ruler 1 (at the back of the skull) in mm (also see photographs below). Record your data in the first column labeled “H”. Skull height measurement: view from the side for two different species – note: upright ruler in the front of skull is removed. Skull Measurement Guide W – Width of the brain case: Position 3 rulers as shown in the photographs below. Rulers 1 and 2 should stand perpendicular to the table and flat against the skull (imagine 2 rulers flat against where the ears would be positioned). -

Artifacts and Commingled Skeletal Remains from a Well on the Medical College of Virginia Campus: Appendices Douglas W
Virginia Commonwealth University VCU Scholars Compass Office of the President Documents VCU University Archives 6-18-2012 Artifacts and Commingled Skeletal Remains from a Well on the Medical College of Virginia Campus: Appendices Douglas W. Owsley Smithsonian Institution Karin Bruwelheide Smithsonian Institution Merry Outlaw Archaeological and Cultural Solutions Richard L. Jantz University of Tennessee-Knoxville Follow this and additional works at: http://scholarscompass.vcu.edu/arch001 Downloaded from http://scholarscompass.vcu.edu/arch001/5 This Research Report is brought to you for free and open access by the VCU University Archives at VCU Scholars Compass. It has been accepted for inclusion in Office of the President Documents by an authorized administrator of VCU Scholars Compass. For more information, please contact [email protected]. Appendix I. Artifact Catalog C o Site Art Manf Form u Ware Material Color Remarks ID Id Tech n t Flat, everted lip, rolled in; pinched in neck; cylindrical body; slightly 44HE Mouth 001 Jar 28 Glass Colorless domed base with rough 814 Blown pontil on exterior; base dia.: 5 3/4"; 28 fragments Translucent white fabric; glossy white glaze; interior underglaze 44HE Ch Blue and Wheel- 002 Saucer 1 Porcelain cobalt blue decoration 814 Porcelain White Thrown consisting of hanging dots above diaper motif; ca. 1785-1820 Translucent white fabric; colorless glaze; interior hand painted overglaze 44HE Ch Wheel- decoration consisting of 003 Bowl 1 Porcelain Polychrome 814 Porcelain Thrown horizontal lines of black -

Level I to III Craniofacial Approaches Based on Barrow Classification For
Neurosurg Focus 30 (5):E5, 2011 Level I to III craniofacial approaches based on Barrow classification for treatment of skull base meningiomas: surgical technique, microsurgical anatomy, and case illustrations EMEL AVCı, M.D.,1 ERINÇ AKTÜRE, M.D.,1 HAKAN SEÇKIN, M.D., PH.D.,1 KUTLUAY ULUÇ, M.D.,1 ANDREW M. BAUER, M.D.,1 YUSUF IZCI, M.D.,1 JACQUes J. MORCOS, M.D.,2 AND MUSTAFA K. BAşKAYA, M.D.1 1Department of Neurological Surgery, University of Wisconsin–Madison, Wisconsin; and 2Department of Neurological Surgery, University of Miami, Florida Object. Although craniofacial approaches to the midline skull base have been defined and surgical results have been published, clear descriptions of these complex approaches in a step-wise manner are lacking. The objective of this study is to demonstrate the surgical technique of craniofacial approaches based on Barrow classification (Levels I–III) and to study the microsurgical anatomy pertinent to these complex craniofacial approaches. Methods. Ten adult cadaveric heads perfused with colored silicone and 24 dry human skulls were used to study the microsurgical anatomy and to demonstrate craniofacial approaches in a step-wise manner. In addition to cadaveric studies, case illustrations of anterior skull base meningiomas were presented to demonstrate the clinical application of the first 3 (Levels I–III) approaches. Results. Cadaveric head dissection was performed in 10 heads using craniofacial approaches. Ethmoid and sphe- noid sinuses, cribriform plate, orbit, planum sphenoidale, clivus, sellar, and parasellar regions were shown at Levels I, II, and III. In 24 human dry skulls (48 sides), a supraorbital notch (85.4%) was observed more frequently than the supraorbital foramen (14.6%). -

The Frontal Bone As a Proxy for Sex Estimation in Humans: a Geometric
Louisiana State University LSU Digital Commons LSU Master's Theses Graduate School 2014 The frontal bone as a proxy for sex estimation in humans: a geometric morphometric analysis Lucy Ann Edwards Hochstein Louisiana State University and Agricultural and Mechanical College, [email protected] Follow this and additional works at: https://digitalcommons.lsu.edu/gradschool_theses Part of the Social and Behavioral Sciences Commons Recommended Citation Hochstein, Lucy Ann Edwards, "The frontal bone as a proxy for sex estimation in humans: a geometric morphometric analysis" (2014). LSU Master's Theses. 1749. https://digitalcommons.lsu.edu/gradschool_theses/1749 This Thesis is brought to you for free and open access by the Graduate School at LSU Digital Commons. It has been accepted for inclusion in LSU Master's Theses by an authorized graduate school editor of LSU Digital Commons. For more information, please contact [email protected]. THE FRONTAL BONE AS A PROXY FOR SEX ESTIMATION IN HUMANS: A GEOMETRIC MORPHOMETRIC ANALYSIS A Thesis Submitted to the Graduate Faculty of the Louisiana State University and Agricultural and Mechanical College in partial fulfillment of the requirements for the degree of Master of Anthropology in The Department of Geography and Anthropology By Lucy A. E. Hochstein B.A., George Mason University, 2009 May 2014 ACKNOWLEDGEMENTS Completing a master’s thesis was the most terrifying aspect of graduate school and I must acknowledge the people and pets that helped me on this adventure. I could not have asked for a better committee chair than Dr. Ginny Listi, who stuck by me when everything fell apart and was always willing to offer support and help me find solutions. -

Supernumerary Bones in the Walls of the Bony Orbit K.Y
Research Article Supernumerary Bones in the Walls of the Bony Orbit K.Y. Manjunath Department of Anatomy, Annapoorna Medical College, NH-47, Shankari Main Road, Periyaseeragapadi Post, Salem-636308 (Received July , 2012) (Accepted January, 2013) Abstract: Occurrence of supernumerary bones in the walls of the orbit especially in the medial wall and the roof has been described in the literature. Studies of the prevalence of supernumerary bones in the bony wall of the orbit are scarce in the literature. Present study was undertaken to find the prevalence of supernumerary bones in the walls of the orbit in a collection of adult Indian skulls. In the present study three hundred and twenty six orbital walls from one hundred and sixty three skulls were examined for the presence of the sutural bones. Their location with reference to the sutures in the walls of the orbit and their size was noted. The supernumerary bones were found in 25 skulls (15.34 %) mainly in the lateral wall (11.04 %) and the roof of the orbit (4.29 %). Prevalence of such supernumerary bones in the walls of the bony orbit is of anthropological interest. Many of the bony ossicles were of sufficiently large size, enough to be visualized on lateral skull X-ray and could easily be mistaken for fracture of the bony wall of the orbit. Key Words: Bony ossicles; Cranium; Orbit; Supernumerary bones; Wormian bones. Introduction: of adult Indian skulls available in the Department of Small islands of bones called the sutural bones Anatomy, St John’s Medical College, Bangalore. are known to occur along the cranial sutures or at the In each skull, the inner surface of the walls of junction of cranial sutures on the cranial vault (Inkster, the bony orbit were inspected carefully for the presence 1951; Black, 2008). -

Eyebrow Peak Position Redefined © 2010 the American Society for Aesthetic Plastic Surgery, Inc
Facial Surgery Aesthetic Surgery Journal 30(3) 297 –300 Eyebrow Peak Position Redefined © 2010 The American Society for Aesthetic Plastic Surgery, Inc. Reprints and permission: http://www .sagepub.com/ journalsPermissions.nav DOI: 10.1177/1090820X10369918 Sylvia Pham; Bradon Wilhelmi, MD; and Arian Mowlavi, MD www.aestheticsurgeryjournal.com Abstract Background: The aesthetically appealing eyebrow shape has been defined by its arch, located near the junction between the medial two-thirds and lateral one-third. The position of this arch has been historically described by arbitrary anatomical landmarks that have no logical structural relationship. Moreover, selection of endoscopic brow lift incision sites that define vector of pull and fixation points have been variably described. Objectives: The authors examine the position of the deep temporal fusion line to determine whether it can act as a more accurate and functional landmark than prior anatomical landmarks for the eyebrow peak position. Methods: Eyebrows were measured in 50 subjects from the medial aspect of the eyebrow to the a) deep temporal fusion line (ridge), b) eyebrow peak (arch), c) lateral aspect of the brow, and d) lateral limbus. Pearson's correlation, descriptive statistics, and student's t test results were obtained. Results: Eyebrow measurements demonstrated that the deep temporal fusion line is the most precise indicator of brow peak position among all examined landmarks. The Pearson correlation value was strongest between brow peak and deep temporal fusion line (P = .860) and a t test confirmed this observation with no significant difference between brow peak and deep temporal fusion line. The lateral limbus and medial two-thirds lateral one- third junction more accurately predict brow peak in females, but the deep temporal fusion line is an equally reliable predictor of brow peak for males and females.