A Description of the Geological Context, Discrete Traits, and Linear Morphometrics of the Middle Pleistocene Hominin from Dali, Shaanxi Province, China
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Dental Pathology, Wear and Developmental Defects in South African Hominins
Dental pathology, wear and developmental defects in South African hominins IAN EDWARD TOWLE A thesis submitted in partial fulfilment of the requirements of Liverpool John Moores University for the degree of Doctor of Philosophy June 2017 Abstract Studying different types of dental pathology, wear, and developmental defects can allow inferences into diet and behaviour in a variety of ways. In this project data on these different variables were collected for South African hominins and compared with extant primates. The species studied include Paranthropus robustus, Australopithecus africanus, A. sediba, early Homo, Homo naledi, baboons, chimpanzees and gorillas. Macroscopic examination of each specimen was performed, with a 10X hand lens used to verify certain pathologies. Variables recorded include antemortem chipping, enamel hypoplasia, caries, occlusal wear, tertiary dentine, abscesses, and periodontal disease. Clear differences in frequencies were found in the different South African hominin species. Homo naledi displays high rates of chipping, especially small fractures above molar wear facets, likely reflecting a diet containing high levels of contaminants. Other noteworthy results include the high levels of pitting enamel hypoplasia in P. robustus molars compared to other species, likely due to a species-specific enamel formation property or developmental disturbance. The low rates of chipping in P. robustus does not fit with this species being a hard food specialist. Instead, the wear best supports a diet of low-quality tough vegetation. Australopithecus africanus likely had a broad diet, with angled molar wear, lack of caries, and high chipping frequencies supporting this conclusion. Seven new carious lesions are described, two from H. naledi and five P. -

Hands-On Human Evolution: a Laboratory Based Approach
Hands-on Human Evolution: A Laboratory Based Approach Developed by Margarita Hernandez Center for Precollegiate Education and Training Author: Margarita Hernandez Curriculum Team: Julie Bokor, Sven Engling A huge thank you to….. Contents: 4. Author’s note 5. Introduction 6. Tips about the curriculum 8. Lesson Summaries 9. Lesson Sequencing Guide 10. Vocabulary 11. Next Generation Sunshine State Standards- Science 12. Background information 13. Lessons 122. Resources 123. Content Assessment 129. Content Area Expert Evaluation 131. Teacher Feedback Form 134. Student Feedback Form Lesson 1: Hominid Evolution Lab 19. Lesson 1 . Student Lab Pages . Student Lab Key . Human Evolution Phylogeny . Lab Station Numbers . Skeletal Pictures Lesson 2: Chromosomal Comparison Lab 48. Lesson 2 . Student Activity Pages . Student Lab Key Lesson 3: Naledi Jigsaw 77. Lesson 3 Author’s note Introduction Page The validity and importance of the theory of biological evolution runs strong throughout the topic of biology. Evolution serves as a foundation to many biological concepts by tying together the different tenants of biology, like ecology, anatomy, genetics, zoology, and taxonomy. It is for this reason that evolution plays a prominent role in the state and national standards and deserves thorough coverage in a classroom. A prime example of evolution can be seen in our own ancestral history, and this unit provides students with an excellent opportunity to consider the multiple lines of evidence that support hominid evolution. By allowing students the chance to uncover the supporting evidence for evolution themselves, they discover the ways the theory of evolution is supported by multiple sources. It is our hope that the opportunity to handle our ancestors’ bone casts and examine real molecular data, in an inquiry based environment, will pique the interest of students, ultimately leading them to conclude that the evidence they have gathered thoroughly supports the theory of evolution. -

Is the Skeleton Male Or Female? the Pelvis Tells the Story
Activity: Is the Skeleton Male or Female? The pelvis tells the story. Distinct features adapted for childbearing distinguish adult females from males. Other bones and the skull also have features that can indicate sex, though less reliably. In young children, these sex-related features are less obvious and more difficult to interpret. Subtle sex differences are detectable in younger skeletons, but they become more defined following puberty and sexual maturation. What are the differences? Compare the two illustrations below in Figure 1. Female Pelvic Bones Male Pelvic Bones Broader sciatic notch Narrower sciatic notch Raised auricular surface Flat auricular surface Figure 1. Female and male pelvic bones. (Source: Smithsonian Institution, illustrated by Diana Marques) Figure 2. Pelvic bone of the skeleton in the cellar. (Source: Smithsonian Institution) Skull (Cranium and Mandible) Male Skulls Generally larger than female Larger projections behind the Larger brow ridges, with sloping, ears (mastoid processes) less rounded forehead Square chin with a more vertical Greater definition of muscle (acute) angle of the jaw attachment areas on the back of the head Figure 3. Male skulls. (Source: Smithsonian Institution, illustrated by Diana Marques) Female Skulls Smoother bone surfaces where Smaller projections behind the muscles attach ears (mastoid processes) Less pronounced brow ridges, Chin more pointed, with a larger, with more vertical forehead obtuse angle of the jaw Sharp upper margins of the eye orbits Figure 4. Female skulls. (Source: Smithsonian Institution, illustrated by Diana Marques) What Do You Think? Comparing the skull from the cellar in Figure 5 (below) with the illustrated male and female skulls in Figures 3 and 4, write Male or Female to note the sex depicted by each feature. -

Morfofunctional Structure of the Skull
N.L. Svintsytska V.H. Hryn Morfofunctional structure of the skull Study guide Poltava 2016 Ministry of Public Health of Ukraine Public Institution «Central Methodological Office for Higher Medical Education of MPH of Ukraine» Higher State Educational Establishment of Ukraine «Ukranian Medical Stomatological Academy» N.L. Svintsytska, V.H. Hryn Morfofunctional structure of the skull Study guide Poltava 2016 2 LBC 28.706 UDC 611.714/716 S 24 «Recommended by the Ministry of Health of Ukraine as textbook for English- speaking students of higher educational institutions of the MPH of Ukraine» (minutes of the meeting of the Commission for the organization of training and methodical literature for the persons enrolled in higher medical (pharmaceutical) educational establishments of postgraduate education MPH of Ukraine, from 02.06.2016 №2). Letter of the MPH of Ukraine of 11.07.2016 № 08.01-30/17321 Composed by: N.L. Svintsytska, Associate Professor at the Department of Human Anatomy of Higher State Educational Establishment of Ukraine «Ukrainian Medical Stomatological Academy», PhD in Medicine, Associate Professor V.H. Hryn, Associate Professor at the Department of Human Anatomy of Higher State Educational Establishment of Ukraine «Ukrainian Medical Stomatological Academy», PhD in Medicine, Associate Professor This textbook is intended for undergraduate, postgraduate students and continuing education of health care professionals in a variety of clinical disciplines (medicine, pediatrics, dentistry) as it includes the basic concepts of human anatomy of the skull in adults and newborns. Rewiewed by: O.M. Slobodian, Head of the Department of Anatomy, Topographic Anatomy and Operative Surgery of Higher State Educational Establishment of Ukraine «Bukovinian State Medical University», Doctor of Medical Sciences, Professor M.V. -

Introduction to the Skeletal System and the Axial Skeleton 155
chapter Introduction to the 7 Skeletal System and the Axial Skeleton CHAPTER OVERVIEW OBJECTIVES 7.1 Introduction to the Skeletal System ……………… 153 1. Describe the gross anatomy and structure of a long 7.2 Bone Structure ………………………………………… 154 bone 7.3 Bone Histology ………………………………………… 155 2. Describe and compare the underlying histology of spongy and compact bone. 7.4 The Human Skeleton: Axial and Appendicular Divisions …………………………………………………… 156 3. List the five general shapes of bones. 7.5 Bone Classification and Markings ………………… 157 4. Describe and compare the different kinds of bone markings visible on the skeleton. 7.6 Axial Skeleton …………………………………………… 159 7.6a Cranium 5. Identify the components of the axial skeleton: cranial, 7.6b Facial facial, hyoid, vertebra, ribs and sternum. 7.6c Hyoid Bone 7.6d Vertebral Column 7.6e Thoracic Cage 7.1 Introduction to the Skeletal System The skeletal system serves to support the body’s soft tissues and to protect the body’s soft internal organs. Another important function that the bones have is to store materials such as calcium, phosphorus and lipids. Additionally, blood cells are synthesized in the red bone marrow to be released into the bloodstream. Bones serve as levers for the muscular system, working with them to produce movement and maintain posture. The human body contains 2 major kinds of bone tissue: compact and spongy. Compact bone (dense bone) is found on the outer surface of bones and serves as a place to absorb most of the stress on the bones. Spongy bone (cancellous tissue) is found on the inside of the compact bone layer. -

Download Poster
Sutural Variability in the Hominoid Anterior Cranial Fossa Robert McCarthy, Monica Kunkel, Department of Biological Sciences, Benedictine University Email contact information: [email protected]; [email protected] SAMPLE ABSTRACT Table 2. Results split by age. Table 4. Specimens used in scaling analyses. Group Age A-M434; M555 This study Combined In anthropoids, the orbital plates of frontal bone meet at a “retro-ethmoid” Species Symbol FMNH CMNH USNM % Contact S-E S-E S-E % % % frontal suture in the midline anterior cranial fossa (ACF), separating the contact/n contact/n contact/n presphenoid and mesethmoid bones. Previous research indicates that this Hylobatid 0 0 23 13.0 Hylobatid Adult 0/6 0 4/13 30.8 4/19 21.1 configuration appears variably in chimpanzees and gorillas and infrequently in modern humans, with speculation that its incidence is related to differential Juvenile 0/2 0 3/10 30.0 3/12 25.0 Orangutan 6 0 37 100.0 growth of the brain and orbits, size of the brow ridges or face, or upper facial Combined 0/9 0 7/23 30.4 7/32 21.9 prognathism. We collected qualitative and quantitative data from 164 Gorilla 3 0 4 28.6 previously-opened cranial specimens from 15 hominoid species in addition to A Orangutan Adult 12/12 100.0 21/21 100.0 21/21 100.0 Chimpanzee 1 0 7 57.1 qualitative observations on non-hominoid and hominin specimens in order to Juvenile 13/13 100.0 13/13 100.0 13/13 100.0 (1) document sutural variability in the primate ACF, (2) rethink the evolutionary trajectory of frontal bone contribution to the midline ACF, and Combined 26/26 100.0 34/34 100.0 34/34 34/34 Human 0 83 0 91.8 (3) create a database of ACF observations and measurements which can be Gorilla Adult 3/7 42.9 1/2 50.0 3/7 42.9 used to test hypotheses about structural relationships in the hominoid ACF. -

Topographical Anatomy and Morphometry of the Temporal Bone of the Macaque
Folia Morphol. Vol. 68, No. 1, pp. 13–22 Copyright © 2009 Via Medica O R I G I N A L A R T I C L E ISSN 0015–5659 www.fm.viamedica.pl Topographical anatomy and morphometry of the temporal bone of the macaque J. Wysocki 1Clinic of Otolaryngology and Rehabilitation, II Medical Faculty, Warsaw Medical University, Poland, Kajetany, Nadarzyn, Poland 2Laboratory of Clinical Anatomy of the Head and Neck, Institute of Physiology and Pathology of Hearing, Poland, Kajetany, Nadarzyn, Poland [Received 7 July 2008; Accepted 10 October 2008] Based on the dissections of 24 bones of 12 macaques (Macaca mulatta), a systematic anatomical description was made and measurements of the cho- sen size parameters of the temporal bone as well as the skull were taken. Although there is a small mastoid process, the general arrangement of the macaque’s temporal bone structures is very close to that which is observed in humans. The main differences are a different model of pneumatisation and the presence of subarcuate fossa, which possesses considerable dimensions. The main air space in the middle ear is the mesotympanum, but there are also additional air cells: the epitympanic recess containing the head of malleus and body of incus, the mastoid cavity, and several air spaces on the floor of the tympanic cavity. The vicinity of the carotid canal is also very well pneuma- tised and the walls of the canal are very thin. The semicircular canals are relatively small, very regular in shape, and characterized by almost the same dimensions. The bony walls of the labyrinth are relatively thin. -

Factors Influencing the Articular Eminence of the Temporomandibular Joint (Review)
EXPERIMENTAL AND THERAPEUTIC MEDICINE 22: 1084, 2021 Factors influencing the articular eminence of the temporomandibular joint (Review) MARIA JUSTINA ROXANA VÎRLAN1, DIANA LORETA PĂUN2, ELENA NICOLETA BORDEA3, ANGELO PELLEGRINI3, ARSENIE DAN SPÎNU4, ROXANA VICTORIA IVAȘCU5, VICTOR NIMIGEAN5 and VANDA ROXANA NIMIGEAN1 1Discipline of Oral Rehabilitation, Faculty of Dental Medicine; 2Discipline of Endocrinology, Faculty of Medicine; 3Department of Specific Disciplines, Faculty of Midwifery and Nursing;4 Discipline of Urology, ‘Dr Carol Davila’ Central Military Emergency University Hospital, Faculty of Medicine; 5Discipline of Anatomy, Faculty of Dental Medicine, ‘Carol Davila’ University of Medicine and Pharmacy, 020021 Bucharest, Romania Received April 28, 2021; Accepted May 28, 2021 DOI: 10.3892/etm.2021.10518 Abstract. The temporomandibular joint (TMJ), the most 4. Factors influencing the inclination of the articular eminence complex and evolved joint in humans, presents two articular 5. Biological sex surfaces: the condyle of the mandible and the articular eminence 6. Age (AE) of the temporal bone. AE is the anterior root of the zygo‑ 7. Edentulism matic process of the temporal bone and has an anterior and 8. Conclusions a posterior slope, the latter being also known as the articular surface. AE is utterly important in the biomechanics of the TMJ, as the mandibular condyle slides along the posterior slope 1. Introduction of the AE while the mandible moves. The aim of this review was to assess significant factors influencing the inclination of The temporomandibular joint (TMJ), the most complex and the AE, especially modifications caused by aging, biological evolved joint in humans, is defined by the paired joints that sex or edentulism. -
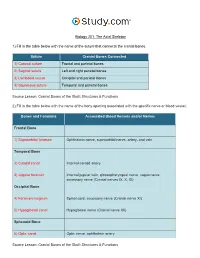
The Axial Skeleton Visual Worksheet
Biology 201: The Axial Skeleton 1) Fill in the table below with the name of the suture that connects the cranial bones. Suture Cranial Bones Connected 1) Coronal suture Frontal and parietal bones 2) Sagittal suture Left and right parietal bones 3) Lambdoid suture Occipital and parietal bones 4) Squamous suture Temporal and parietal bones Source Lesson: Cranial Bones of the Skull: Structures & Functions 2) Fill in the table below with the name of the bony opening associated with the specific nerve or blood vessel. Bones and Foramina Associated Blood Vessels and/or Nerves Frontal Bone 1) Supraorbital foramen Ophthalmic nerve, supraorbital nerve, artery, and vein Temporal Bone 2) Carotid canal Internal carotid artery 3) Jugular foramen Internal jugular vein, glossopharyngeal nerve, vagus nerve, accessory nerve (Cranial nerves IX, X, XI) Occipital Bone 4) Foramen magnum Spinal cord, accessory nerve (Cranial nerve XI) 5) Hypoglossal canal Hypoglossal nerve (Cranial nerve XII) Sphenoid Bone 6) Optic canal Optic nerve, ophthalmic artery Source Lesson: Cranial Bones of the Skull: Structures & Functions 3) Label the anterior view of the skull below with its correct feature. Frontal bone Palatine bone Ethmoid bone Nasal septum: Perpendicular plate of ethmoid bone Sphenoid bone Inferior orbital fissure Inferior nasal concha Maxilla Orbit Vomer bone Supraorbital margin Alveolar process of maxilla Middle nasal concha Inferior nasal concha Coronal suture Mandible Glabella Mental foramen Nasal bone Parietal bone Supraorbital foramen Orbital canal Temporal bone Lacrimal bone Orbit Alveolar process of mandible Superior orbital fissure Zygomatic bone Infraorbital foramen Source Lesson: Facial Bones of the Skull: Structures & Functions 4) Label the right lateral view of the skull below with its correct feature. -
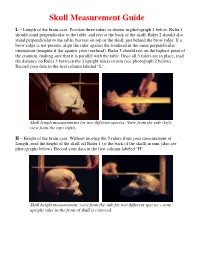
AP Skull Measurement Guide, Laminate
Skull Measurement Guide L – Length of the brain case: Position three rulers as shown in photograph 1 below. Ruler 1 should stand perpendicular to the table, and rest at the back of the skull. Ruler 2 should also stand perpendicular to the table, but rest on top of the skull, just behind the brow ridge. If a brow ridge is not present, align the ruler against the forehead in the same perpendicular orientation (imagine it flat against your forehead). Ruler 3 should rest on the highest point of the cranium, making sure that it is parallel with the table. Once all 3 rulers are in place, read the distance on Ruler 3 between the 2 upright rulers in mm (see photograph 2 below). Record your data in the first column labeled “L”. Skull length measurements for two different species: View from the side (left), view from the top (right). H – Height of the brain case: Without moving the 3 rulers from your measurement of Length, read the height of the skull off Ruler 1 (at the back of the skull) in mm (also see photographs below). Record your data in the first column labeled “H”. Skull height measurement: view from the side for two different species – note: upright ruler in the front of skull is removed. Skull Measurement Guide W – Width of the brain case: Position 3 rulers as shown in the photographs below. Rulers 1 and 2 should stand perpendicular to the table and flat against the skull (imagine 2 rulers flat against where the ears would be positioned). -

Artifacts and Commingled Skeletal Remains from a Well on the Medical College of Virginia Campus: Appendices Douglas W
Virginia Commonwealth University VCU Scholars Compass Office of the President Documents VCU University Archives 6-18-2012 Artifacts and Commingled Skeletal Remains from a Well on the Medical College of Virginia Campus: Appendices Douglas W. Owsley Smithsonian Institution Karin Bruwelheide Smithsonian Institution Merry Outlaw Archaeological and Cultural Solutions Richard L. Jantz University of Tennessee-Knoxville Follow this and additional works at: http://scholarscompass.vcu.edu/arch001 Downloaded from http://scholarscompass.vcu.edu/arch001/5 This Research Report is brought to you for free and open access by the VCU University Archives at VCU Scholars Compass. It has been accepted for inclusion in Office of the President Documents by an authorized administrator of VCU Scholars Compass. For more information, please contact [email protected]. Appendix I. Artifact Catalog C o Site Art Manf Form u Ware Material Color Remarks ID Id Tech n t Flat, everted lip, rolled in; pinched in neck; cylindrical body; slightly 44HE Mouth 001 Jar 28 Glass Colorless domed base with rough 814 Blown pontil on exterior; base dia.: 5 3/4"; 28 fragments Translucent white fabric; glossy white glaze; interior underglaze 44HE Ch Blue and Wheel- 002 Saucer 1 Porcelain cobalt blue decoration 814 Porcelain White Thrown consisting of hanging dots above diaper motif; ca. 1785-1820 Translucent white fabric; colorless glaze; interior hand painted overglaze 44HE Ch Wheel- decoration consisting of 003 Bowl 1 Porcelain Polychrome 814 Porcelain Thrown horizontal lines of black -

The Temporomandibular Joint: Pneumatic Temporal Cells Open Into the Articular and Extradural Spaces C
CORE Metadata, citation and similar papers at core.ac.uk Provided by Via Medica Journals Folia Morphol. Vol. 78, No. 3, pp. 630–636 DOI: 10.5603/FM.a2018.0111 O R I G I N A L A R T I C L E Copyright © 2019 Via Medica ISSN 0015–5659 journals.viamedica.pl The temporomandibular joint: pneumatic temporal cells open into the articular and extradural spaces C. Bichir, M.C. Rusu, A.D. Vrapciu, N. Măru “Carol Davila” University of Medicine and Pharmacy of Bucharest, Romania [Received: 1 October 2018; Accepted: 22 November 2018] The pneumatisation of the articular tubercle (PAT) of the temporal squama is a rare condition that modifies the barrier between the temporomandibular joint (TMJ) space and the middle cranial fossa. During a routine examination of the cone-beam computed tomography (CBCT) files of patients who were scanned for dental medical purposes, we identified a case with multiple rare anatomic variations. First, the petrous apex was bilaterally pneumatised. Moreover, bilateral and multilocular PAT were observed, while on one side it was further found that the pneumatic cells were equally dehiscent towards the extradural space and the superior joint space. To the best of our knowledge, such dehiscence has not pre- viously been reported. The two temporomastoid pneumatisations were extended with occipital pneumatisations of the lateral masses and occipital condyles, the latter being an extremely rare evidence. The internal dehiscence of the mandibular canal in the right ramus of the mandible was also noted. Additionally, double mental foramen and impacted third molars were found on the left side.