Response of Acacia Tortilis to Elephant Browsing in Tarangire National Park, Tanzania: Possible Above-Ground Compensation?
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

American Journal of Climate Change, 2018, 7, 5-26 ISSN Online: 2167-9509 ISSN Print: 2167-9495
American Journal of Climate Change, 2018, 7, 5-26 http://www.scirp.org/journal/ajcc ISSN Online: 2167-9509 ISSN Print: 2167-9495 Potential Impacts of Temperature Projections on Selected Large Herbivores in Savanna Ecosystem of Kenya Mildred M. Aduma1*, Gilbert Ouma1, Mohammed Y. Said2, Gordon O. Wayumba3, Philip A. Omondi1, Lucy W. Njino4 1Institute of Climate Change and Adaptation, University of Nairobi, Nairobi, Kenya 2Center for Sustainable Drylands Ecosystems and Societies, University of Nairobi, Nairobi, Kenya 3School of Surveying and Geospatial Sciences, Technical University of Kenya, Nairobi, Kenya 4Directorate of Resource Surveys and Remote Sensing, Nairobi, Kenya How to cite this paper: Aduma, M.M., Abstract Ouma, G., Said, M.Y., Wayumba, G.O., Omondi, P.A. and Njino, L.W. (2018) Po- Due to global land surface warming, severe temperature events are expected to tential Impacts of Temperature Projections occur more frequently and more extremely causing changes in biodiversity on Selected Large Herbivores in Savanna and altering movement and survival of large herbivores. There are increasing Ecosystem of Kenya. American Journal of Climate Change, 7, 5-26. observations of escalating wildlife range losses worldwide. In this study, we https://doi.org/10.4236/ajcc.2018.71003 investigated 15 large wild herbivores (4 migratory, 1 dispersing and 10 resi- dents) and their potential range changes in relation to projected temperatures Received: November 17, 2017 Accepted: February 9, 2018 changes based on three Representative Concentration Pathways (RCPs) 2.6, Published: February 12, 2018 4.5 and 8.5. Previous studies of Kenyan savannah have shown that increases in temperature can reduce the densities of wildlife significantly and after certain Copyright © 2018 by authors and thresholds the species can be lost in those landscapes. -

My Somali Book, a Record of Two Shooting Trips
10 MY SOMALI BOOK to learn new things, and wonderfully bright and in- telligent. He is untiring on the march, often a reckless hunter, and will stand b}^ his master splendidly." And again, " I have made many jmigle trips in India and elsewhere, yet in no country have I had such obedient and cheerful followers and such pleasant native companions, despite their faults, as in Somali- land." This estimate is every bit as true as the other. The Somali of the interior is not, as a rule, dishonest, though he can be an exasperating liar on occasion. And he is usualty decent in his dress. Naturally indolent, he prefers to see the women do the work, but he has plenty of energy when he chooses to give it exercise, and in this respect is seen at his best on a shooting trip. And there is no doubt of his courage. A Mahomedan of the Shafai Sect, the Somali is sometimes very particular about his religious observances, but the Arab despises him as not a true Mahomedan at all. There is no occasion to detail the principal tribes, which are split up into innumerable sub-divisions. But mention must be made of the Midgdns, an outcast tribe of professional hunters. They hunt with bow and poisoned arrow, sometimes with dogs, and are wonderful trackers. As to the provision which the country makes for the hunter : the principal game to be met with in Northern Somaliland comprises Elephant, Black Rhinoceros, Lion, Leopard, Chita (hunting-leopard). MY SOMALI BOOK 11 Warthog, Ostrich, and twelve species of Antelope, to wit, Greater and Lesser Kudu, Or3^x (Beisa), Swa3aie's Hartebeest, Sommering's Gazelle (Somali-^Oi(/), Waller's Gazelle (Gerenuk), Clarke's Gazelle {Dibatag), Speke's and Pelzeln's Gazelles (both Dhero in Somali), Baira, Klipspringer {Alahlt) and Phillips' Dik-dik {Sdkdro). -

Animals of Africa
Silver 49 Bronze 26 Gold 59 Copper 17 Animals of Africa _______________________________________________Diamond 80 PYGMY ANTELOPES Klipspringer Common oribi Haggard oribi Gold 59 Bronze 26 Silver 49 Copper 17 Bronze 26 Silver 49 Gold 61 Copper 17 Diamond 80 Diamond 80 Steenbok 1 234 5 _______________________________________________ _______________________________________________ Cape grysbok BIG CATS LECHWE, KOB, PUKU Sharpe grysbok African lion 1 2 2 2 Common lechwe Livingstone suni African leopard***** Kafue Flats lechwe East African suni African cheetah***** _______________________________________________ Red lechwe Royal antelope SMALL CATS & AFRICAN CIVET Black lechwe Bates pygmy antelope Serval Nile lechwe 1 1 2 2 4 _______________________________________________ Caracal 2 White-eared kob DIK-DIKS African wild cat Uganda kob Salt dik-dik African golden cat CentralAfrican kob Harar dik-dik 1 2 2 African civet _______________________________________________ Western kob (Buffon) Guenther dik-dik HYENAS Puku Kirk dik-dik Spotted hyena 1 1 1 _______________________________________________ Damara dik-dik REEDBUCKS & RHEBOK Brown hyena Phillips dik-dik Common reedbuck _______________________________________________ _______________________________________________African striped hyena Eastern bohor reedbuck BUSH DUIKERS THICK-SKINNED GAME Abyssinian bohor reedbuck Southern bush duiker _______________________________________________African elephant 1 1 1 Sudan bohor reedbuck Angolan bush duiker (closed) 1 122 2 Black rhinoceros** *** Nigerian -

A Scoping Review of Viral Diseases in African Ungulates
veterinary sciences Review A Scoping Review of Viral Diseases in African Ungulates Hendrik Swanepoel 1,2, Jan Crafford 1 and Melvyn Quan 1,* 1 Vectors and Vector-Borne Diseases Research Programme, Department of Veterinary Tropical Disease, Faculty of Veterinary Science, University of Pretoria, Pretoria 0110, South Africa; [email protected] (H.S.); [email protected] (J.C.) 2 Department of Biomedical Sciences, Institute of Tropical Medicine, 2000 Antwerp, Belgium * Correspondence: [email protected]; Tel.: +27-12-529-8142 Abstract: (1) Background: Viral diseases are important as they can cause significant clinical disease in both wild and domestic animals, as well as in humans. They also make up a large proportion of emerging infectious diseases. (2) Methods: A scoping review of peer-reviewed publications was performed and based on the guidelines set out in the Preferred Reporting Items for Systematic Reviews and Meta-Analyses (PRISMA) extension for scoping reviews. (3) Results: The final set of publications consisted of 145 publications. Thirty-two viruses were identified in the publications and 50 African ungulates were reported/diagnosed with viral infections. Eighteen countries had viruses diagnosed in wild ungulates reported in the literature. (4) Conclusions: A comprehensive review identified several areas where little information was available and recommendations were made. It is recommended that governments and research institutions offer more funding to investigate and report viral diseases of greater clinical and zoonotic significance. A further recommendation is for appropriate One Health approaches to be adopted for investigating, controlling, managing and preventing diseases. Diseases which may threaten the conservation of certain wildlife species also require focused attention. -

List of 28 Orders, 129 Families, 598 Genera and 1121 Species in Mammal Images Library 31 December 2013
What the American Society of Mammalogists has in the images library LIST OF 28 ORDERS, 129 FAMILIES, 598 GENERA AND 1121 SPECIES IN MAMMAL IMAGES LIBRARY 31 DECEMBER 2013 AFROSORICIDA (5 genera, 5 species) – golden moles and tenrecs CHRYSOCHLORIDAE - golden moles Chrysospalax villosus - Rough-haired Golden Mole TENRECIDAE - tenrecs 1. Echinops telfairi - Lesser Hedgehog Tenrec 2. Hemicentetes semispinosus – Lowland Streaked Tenrec 3. Microgale dobsoni - Dobson’s Shrew Tenrec 4. Tenrec ecaudatus – Tailless Tenrec ARTIODACTYLA (83 genera, 142 species) – paraxonic (mostly even-toed) ungulates ANTILOCAPRIDAE - pronghorns Antilocapra americana - Pronghorn BOVIDAE (46 genera) - cattle, sheep, goats, and antelopes 1. Addax nasomaculatus - Addax 2. Aepyceros melampus - Impala 3. Alcelaphus buselaphus - Hartebeest 4. Alcelaphus caama – Red Hartebeest 5. Ammotragus lervia - Barbary Sheep 6. Antidorcas marsupialis - Springbok 7. Antilope cervicapra – Blackbuck 8. Beatragus hunter – Hunter’s Hartebeest 9. Bison bison - American Bison 10. Bison bonasus - European Bison 11. Bos frontalis - Gaur 12. Bos javanicus - Banteng 13. Bos taurus -Auroch 14. Boselaphus tragocamelus - Nilgai 15. Bubalus bubalis - Water Buffalo 16. Bubalus depressicornis - Anoa 17. Bubalus quarlesi - Mountain Anoa 18. Budorcas taxicolor - Takin 19. Capra caucasica - Tur 20. Capra falconeri - Markhor 21. Capra hircus - Goat 22. Capra nubiana – Nubian Ibex 23. Capra pyrenaica – Spanish Ibex 24. Capricornis crispus – Japanese Serow 25. Cephalophus jentinki - Jentink's Duiker 26. Cephalophus natalensis – Red Duiker 1 What the American Society of Mammalogists has in the images library 27. Cephalophus niger – Black Duiker 28. Cephalophus rufilatus – Red-flanked Duiker 29. Cephalophus silvicultor - Yellow-backed Duiker 30. Cephalophus zebra - Zebra Duiker 31. Connochaetes gnou - Black Wildebeest 32. Connochaetes taurinus - Blue Wildebeest 33. Damaliscus korrigum – Topi 34. -
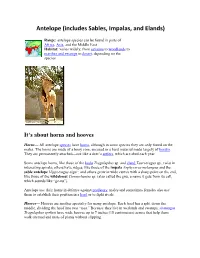
Antelope (Includes Sables, Impalas, and Elands)
Antelope (includes Sables, Impalas, and Elands) Range: antelope species can be found in parts of Africa, Asia, and the Middle East Habitat: varies widely, from savanna to woodlands to marshes and swamps to desert, depending on the species It’s about horns and hooves Horns— All antelope species have horns, although in some species they are only found on the males. The horns are made of a bony core, encased in a hard material made largely of keratin. They are permanently attached—not like a deer’s antlers, which are shed each year. Some antelope horns, like those of the kudu Tragelaphus sp. and eland Taurotragus sp., twist in interesting spirals; others have ridges, like those of the impala Aephyceros melampus and the sable antelope Hippotragus niger; and others grow in wide curves with a sharp point on the end, like those of the wildebeest Connochaetes sp. (also called the gnu, a name it gets from its call, which sounds like “ge-nu”). Antelope use their horns in defense against predators; males and sometimes females also use them to establish their position in a herd or to fight rivals Hooves— Hooves are another specialty for many antelope. Each hoof has a split down the middle, dividing the hoof into two “toes.” Because they live in wetlands and swamps, sitatungas Tragelaphus spekeii have wide hooves up to 7 inches (18 centimeters) across that help them walk on mud and mats of plants without slipping. Nile lechwes Kobus magaceros, which also live in swampy areas, have long, pointed hooves to give them sure footing in the water. -

GNUSLETTER Vol 37#2.Pdf
GNUSLETTER Volume 37 / Number 2 December 2020 ANTELOPE SPECIALIST GROUP IUCN Species Survival Commission Antelope Specialist Group GNUSLETTER is the biannual newsletter of the IUCN Species Survival Commission Antelope Specialist Group (ASG). First published in 1982 by first ASG Chair Richard D. Estes, the intent of GNUSLETTER, then and today, is the dissemination of reports and information regarding antelopes and their conservation. ASG Members are an important network of individuals and experts working across disciplines throughout Africa, Asia and America. Contributions (original articles, field notes, other material relevant to antelope biology, ecology, and conservation) are welcomed and should be sent to the editor. Today GNUSLETTER is published in English in electronic form and distributed widely to members and non-members, and to the IUCN SSC global conservation network. To be added to the distribution list please contact [email protected]. GNUSLETTER Editorial Board - David Mallon, ASG Co-Chair - Philippe Chardonnet, ASG Co-Chair ASG Program Office - Tania Gilbert, Marwell Wildife The Antelope Specialist Group Program Office is hosted and supported by Marwell Wildlife https://www.marwell.org.uk The designation of geographical entities in this report does not imply the expression of any opinion on the part of IUCN, the Species Survival Commission, or the Antelope Specialist Group concerning the legal status of any country, territory or area, or concerning the delimitation of any frontiers or boundaries. Views expressed in GNUSLETTER are those of the individual authors, Cover photo: Young female bushbuck (Tragelaphus scriptus), W National Park and Biosphere Reserve, Niger (© Daniel Cornélis) 2 GNUSLETTER Volume 37 Number 2 December 2020 FROM IUCN AND ASG………………………………………………………. -

Digestive Strategies in Ruminants and Nonruminants S.E. Van Wieren
DIGESTIVE STRATEGIES IN RUMINANTS AND NONRUMINANTS S.E. VANWIERE N 0000 0714 3726 Promotoren: Dr. ir. S. Tamminga, buitengewoon hoogleraar op het vakgebied van de veevoeding in het bijzonder de voeding van herkauwers Dr. H.H.T. Prins, hoogleraar in het natuurbeheer in de tropen en oecologie van vertebraten WMoPûi , ZI 9 S.E. van Wieren DIGESTIVE STRATEGIES IN RUMINANTS AND NONRUMINANTS Proefschrift ter verkrijging van de graad van doctor op gezag van de rector magnificus van de Landbouwuniversiteit Wageningen, dr. C.M. Karssen, in het openbaar te verdedigen op dinsdag 3 december 1996 des namiddags te vier uur in de Aula. aiqo?<? T CIP-DATA KONINKLIJKE BIBLIOTHEEK, DEN HAAG Van Wieren, S.E. Digestive strategies in ruminants and nonruminants / S.E. van Wieren. - Thesis Landbouw Universiteit Wageningen. - With réf. - With summary in Dutch. ISBN 90-5485-611-4 Subject headings: digestion / ruminants /nonruminants / feeding ecology cr.-::i n; . •Y.:r Cover and illustrations: Esther van Nie & Marjolein Wiersma STELLINGEN I De veelvuldig aangehangen gedachte dat paarden, wat betreft de dagelijkse opname van metaboliseerbare energie uit laagwaardig voedsel, superieur zijn aan runderen, wordt niet door alle onderzoek ondersteund en blijkt ook niet altijd uit praktijkervaringen. P. Duncan et al. (1990). Oecologia 84:411-418. R. Meydam. Evaluatie begrazing Meyendel. 1996. II Wilde zwijnen die op een dieet van uitsluitend mast leven, krijgen onherroepelijk eiwitgebrek. III Het concept van duurzame ontwikkeling leidt in de praktijk niet zozeer tot beperkingen aan de groei, maar meer tot de groei van de beperkingen. B. Willers. (1994). Conservation Biology (8):1146-1148 . IV De typisch nederlandse gedachte dat de mens verrijkend heeft gewerkt op de natuur, is een gevaarlijke misvatting. -

Title 50 Part 17
U Title 50—Wildlife and Fisheries Ihe information of the reader. In the annual all other appropriate rules in Parts 17. 217 revision and compilation of this title, the through 227. and 402 still apply to that PART 17—ENDANGERED AND following information may be amended species. In addition, there may be other rules THREATENED WILDLIFE AND PLANTS without public notice: the spelling of species' in this title that relate to such wildlife, e.g.. names, historical range, footnotes, references pon-of-entry requirements. It is not intended to certain other applicable portions of this that the references m the "Special rules" title, synonyms, and more current names. In column list all the regulations of the two Subpart B—Lists any of these revised entries, neither the Services which might apply to the species or § 17.11 Endangered and threatened species, as defined in paragraph (b) of this to the regulations of other Federal agencies wildlife. section, nor its status may be changed without or Stale or local governments. (a) The list in this section contains the following the procedures of Part 424 of this (g) The listing of a particular taxon names of all species of wildlife which have title. includes all lower taxonomic units. For been determined by Ihe Services to be (e) The "historic range" indicates the example, the genus H\lobaies (gibbons) is Endangered or Threatened. It also contains known general distribution of the species or listed as Endangered throughout its entire the names of species of wildlife treated as subspecies as reported in the current scientific range (China. -

Endangered Species
DEPARTMENT OF NATURAL RESOURCES 202-1 Chapter NR 27 ENDANGERED SPECIES NR 27.01 Scope and applicability NR 27.05 Permits for transportation of NR 27.02 Definitions endangered species NR 27.03 Department list NR 27.04 Revision of Wisconsin en- NR 27.06 Exceptions to permit require- dangered species list ments History: Chapter NR 27 as it existed on September 30, 1975 was repealed and a new chapter NR 27 was created effective October 1, 1975. NR 27.01 Scope and applicability. This chapter contain rules necessary to implement the Wisconsin Endangered Species Act of 1971 (section 29.415, Wis. Stats.). The rules in this chapter govern the taking, transportation, possession, processing or sale within this state of any fish or wildlife specified by the department's list of endangered fish and wildlife. History: Cr. Register, September, 1975, No. 237, eff. 10-1-75. NR 27.02 Definitions. As used in this chapter: (1) "Department list of endangered species" shall consist of 2 lists: the federal endangered domestic and foreign species and the Wisconsin endangered species. (2) "Federal endangered domestic and foreign species list" shall mean the list of species or subspecies of fish and wildlife native to the United States that are threatened with extinction and species or subspecies of fish and wildlife found in other countries that are threatened with worldwide extinction, as published in the Code of Federal Regulations, Title 50, revised as of January 1, 1971 and as subsequently amended. (3) "Fish and wildlife" shall mean any species or subspecies of any mammal, fish wild bird, amphibian, reptile, mollusk or crustacean, any part, products, eggs or offspring thereof or the dead body or parts thereof. -

The Practical Side of Immunocontraception: Zona Proteins and Wildlife
This article appeared in a journal published by Elsevier. The attached copy is furnished to the author for internal non-commercial research and education use, including for instruction at the authors institution and sharing with colleagues. Other uses, including reproduction and distribution, or selling or licensing copies, or posting to personal, institutional or third party websites are prohibited. In most cases authors are permitted to post their version of the article (e.g. in Word or Tex form) to their personal website or institutional repository. Authors requiring further information regarding Elsevier’s archiving and manuscript policies are encouraged to visit: http://www.elsevier.com/copyright Author's personal copy Journal of Reproductive Immunology 83 (2009) 151–157 Contents lists available at ScienceDirect Journal of Reproductive Immunology journal homepage: www.elsevier.com/locate/jreprimm The practical side of immunocontraception: zona proteins and wildlife J.F. Kirkpatrick a,∗, A. Rowan b, N. Lamberski c, R. Wallace d, K. Frank a, R. Lyda a a The Science and Conservation Center, ZooMontana, Billings, MT 59106, USA b The Humane Society of the United States, Gaithersburg, MD, USA c San Diego Wild Animal Park, Escondido, CA, USA d Milwaukee County Zoo, Milwaukee, WI, USA article info abstract Article history: With shrinking habitat, the humane control of certain wildlife populations is relevant. The Received 31 December 2008 contraceptive vaccine based on native porcine zona pellucida (PZP) has been applied to Received in revised form 16 June 2009 various wildlife populations for 20 years. Prominent efforts include wild horses, urban Accepted 16 June 2009 deer, zoo animals and African elephants, among others. -

Ethiopia 2019
Ethiopia 2019 Mountain Nyala Hunting in With a 100 % success rate and an average of 37”, our partner Northern Operations Ethiopia Africa is extremely proud to say that nobody can compare with the consistency in quality of Mt. Nyala that they have taken in the recent years. They have never taken a Mt. Nyala under 35” and they intend to stay true to this throughout their years of operation!! Their concessions are managed and monitored very seriously, giving us top quality conditions to produce excellent trophies. Arrival / International Flight & Paperwork: Planning your trip to visit Ethiopia is a very simple process. Once your safari has been confirmed, all paperwork for gun permits and hunting licenses will be provided to you 6 months before your scheduled departure. We will provide hands on help with all the information, paperwork, and questions you may have. Detailed information about the agent or PH that will pick you up, will be provided to you 15-30 days before departure. Hotel That night you will overnight and rest in Addis, at the hotel of your choice, either the Sheraton, a 5-star hotel with all amenities, or the Hilton a 4-star hotel. The third choice would be Intercontinental Hotel a 3-star hotel, which is comfortable. Your PH will brief you about all details concerning your next day departure to the hunting cam / area. The Drive to Camp The duration of the drive to camp, varies on what area you will be hunting, however you can expect between a 4-6 hour car rides to camp.