The Braincase of Two Late Cretaceous Asian Multituberculates Studied by Serial Sections
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

MR Imaging of the Orbital Apex
J Korean Radiol Soc 2000;4 :26 9-0 6 1 6 MR Imaging of the Orbital Apex: An a to m y and Pat h o l o g y 1 Ho Kyu Lee, M.D., Chang Jin Kim, M.D.2, Hyosook Ahn, M.D.3, Ji Hoon Shin, M.D., Choong Gon Choi, M.D., Dae Chul Suh, M.D. The apex of the orbit is basically formed by the optic canal, the superior orbital fis- su r e , and their contents. Space-occupying lesions in this area can result in clinical d- eficits caused by compression of the optic nerve or extraocular muscles. Even vas c u l a r changes in the cavernous sinus can produce a direct mass effect and affect the orbit ap e x. When pathologic changes in this region is suspected, contrast-enhanced MR imaging with fat saturation is very useful. According to the anatomic regions from which the lesions arise, they can be classi- fied as belonging to one of five groups; lesions of the optic nerve-sheath complex, of the conal and intraconal spaces, of the extraconal space and bony orbit, of the cav- ernous sinus or diffuse. The characteristic MR findings of various orbital lesions will be described in this paper. Index words : Orbit, diseases Orbit, MR The apex of the orbit is a complex region which con- tains many nerves, vessels, soft tissues, and bony struc- Anatomy of the orbital apex tures such as the superior orbital fissure and the optic canal (1-3), and is likely to be involved in various dis- The orbital apex region consists of the optic nerve- eases (3). -

Dome-Headed, Small-Brained Island Mammal from the Late Cretaceous of Romania
Dome-headed, small-brained island mammal from the Late Cretaceous of Romania Zoltán Csiki-Savaa,1, Mátyás Vremirb, Jin Mengc, Stephen L. Brusatted, and Mark A. Norellc aLaboratory of Paleontology, Faculty of Geology and Geophysics, University of Bucharest, 010041 Bucharest, Romania; bDepartment of Natural Sciences, Transylvanian Museum Society, 400009 Cluj-Napoca, Romania; cDivision of Paleontology, American Museum of Natural History, New York, NY 10024; and dSchool of GeoSciences, Grant Institute, University of Edinburgh, EH9 3FE Edinburgh, United Kingdom Edited by Neil H. Shubin, The University of Chicago, Chicago, IL, and approved March 26, 2018 (received for review January 20, 2018) The island effect is a well-known evolutionary phenomenon, in describe the anatomy of kogaionids in detail, include them in a which island-dwelling species isolated in a resource-limited envi- comprehensive phylogenetic analysis, estimate their body sizes, ronment often modify their size, anatomy, and behaviors compared and present a reconstruction of their brain and sense organs. with mainland relatives. This has been well documented in modern This species exhibits several features that we interpret as re- and Cenozoic mammals, but it remains unclear whether older, more lated to its insular habitat, most notably a brain that is sub- primitive Mesozoic mammals responded in similar ways to island stantially reduced in size compared with close relatives and habitats. We describe a reasonably complete and well-preserved skeleton of a kogaionid, an enigmatic radiation of Cretaceous island- mainland contemporaries, demonstrating that some Mesozoic dwelling multituberculate mammals previously represented by frag- mammals were susceptible to the island effect like in more mentary fossils. -

The Morphometric Study of Occurrence and Variations of Foramen Ovale S
Research Article The morphometric study of occurrence and variations of foramen ovale S. Ajrish George*, M. S. Thenmozhi ABSTRACT Background: Foramen vale is one of the important foramina present in the sphenoid bone. Anatomically it is located in the greater wing of the sphenoid bone. The foramen ovale is situated posterolateral to the foramen rotundum and anteromedial to the foramen spinosum. The foramen spinosum is present posterior to the foramen ovale. The carotid canal is present posterior and medial to the foramen spinosum and the foramen rotundum is present anterior to the foramen ovale. The structures which pass through the foramen ovale are the mandibular nerve, emissary vein, accessory middle meningeal artery, and lesser petrosal nerve. The sphenoid bone has a body, a pair of greater wing, pair of lesser wing, pair of lateral pterygoid plate, and a pair of medial pterygoid plate. Aim: The study involves the assessment of any additional features in foramen ovale in dry South Indian skulls. Materials and Methods: This study involves examination of dry adult skulls. First, the foramen ovale is located, and then it is carefully examined for presence of alterations and additional features, and is recorded following computing the data and analyzing it. Results: The maximum length of foramen ovale on the right and left was 10.1 mm, 4.3 mm, respectively. The minimum length of the foramen in right and left was 9.1 mm, 3.2 mm, respectively. The maximum width of foramen ovale on the right and left was 4.8 mm and 2.3 mm, respectively. The minimum width of the foramen in the right and the left side was 5.7 mm and 2.9 mm, respectively. -

Craniodental Anatomy of a New Late Cretaceous Multituberculate Mammal from Udan Sayr, Mongolia
University of Louisville ThinkIR: The University of Louisville's Institutional Repository Electronic Theses and Dissertations 8-2014 Craniodental anatomy of a new late cretaceous multituberculate mammal from Udan Sayr, Mongolia. Amir Subhash Sheth University of Louisville Follow this and additional works at: https://ir.library.louisville.edu/etd Part of the Anatomy Commons, and the Medical Neurobiology Commons Recommended Citation Sheth, Amir Subhash, "Craniodental anatomy of a new late cretaceous multituberculate mammal from Udan Sayr, Mongolia." (2014). Electronic Theses and Dissertations. Paper 1317. https://doi.org/10.18297/etd/1317 This Master's Thesis is brought to you for free and open access by ThinkIR: The nivU ersity of Louisville's Institutional Repository. It has been accepted for inclusion in Electronic Theses and Dissertations by an authorized administrator of ThinkIR: The nivU ersity of Louisville's Institutional Repository. This title appears here courtesy of the author, who has retained all other copyrights. For more information, please contact [email protected]. CRANIODENTAL ANATOMY OF A NEW LATE CRETACEOUS MULTITUBERCULATE MAMMAL FROM UDAN SAYR, MONGOLIA By Amir Subhash Sheth B.A., Centre College, 2010 A Thesis Submitted to the Faculty of the School of Medicine of the University of Louisville in Partial Fulfillment of the Requirements for the Degree of Master of Science Department of Anatomical Sciences and Neurobiology University of Louisville Louisville, Kentucky August 2014 CRANIODENTAL ANATOMY OF A NEW LATE CRETACEOUS MULTITUBERCULATE MAMMAL FROM UDAN SAYR, MONGOLIA By Amir Subhash Sheth B.A., Centre College, 2010 A Thesis Approved on July 18th, 2014 By the Following Thesis Committee: ________________________________ (Guillermo W. -

Mammals from the Mesozoic of Mongolia
Mammals from the Mesozoic of Mongolia Introduction and Simpson (1926) dcscrihed these as placental (eutherian) insectivores. 'l'he deltathcroids originally Mongolia produces one of the world's most extraordi- included with the insectivores, more recently have narily preserved assemblages of hlesozoic ma~nmals. t)een assigned to the Metatheria (Kielan-Jaworowska Unlike fossils at most Mesozoic sites, Inany of these and Nesov, 1990). For ahout 40 years these were the remains are skulls, and in some cases these are asso- only Mesozoic ~nanimalsknown from Mongolia. ciated with postcranial skeletons. Ry contrast, 'I'he next discoveries in Mongolia were made by the Mesozoic mammals at well-known sites in North Polish-Mongolian Palaeontological Expeditions America and other continents have produced less (1963-1971) initially led by Naydin Dovchin, then by complete material, usually incomplete jaws with den- Rinchen Barsbold on the Mongolian side, and Zofia titions, or isolated teeth. In addition to the rich Kielan-Jaworowska on the Polish side, Kazi~nierz samples of skulls and skeletons representing Late Koualski led the expedition in 1964. Late Cretaceous Cretaceous mam~nals,certain localities in Mongolia ma~nmalswere collected in three Gohi Desert regions: are also known for less well preserved, but important, Bayan Zag (Djadokhta Formation), Nenlegt and remains of Early Cretaceous mammals. The mammals Khulsan in the Nemegt Valley (Baruungoyot from hoth Early and Late Cretaceous intervals have Formation), and llcrmiin 'ISav, south-\vest of the increased our understanding of diversification and Neniegt Valley, in the Red beds of Hermiin 'rsav, morphologic variation in archaic mammals. which have heen regarded as a stratigraphic ecluivalent Potentially this new information has hearing on the of the Baruungoyot Formation (Gradzinslti r't crl., phylogenetic relationships among major branches of 1977). -

Morphology of the Foramen Magnum in Young Eastern European Adults
Folia Morphol. Vol. 71, No. 4, pp. 205–216 Copyright © 2012 Via Medica O R I G I N A L A R T I C L E ISSN 0015–5659 www.fm.viamedica.pl Morphology of the foramen magnum in young Eastern European adults F. Burdan1, 2, J. Szumiło3, J. Walocha4, L. Klepacz5, B. Madej1, W. Dworzański1, R. Klepacz3, A. Dworzańska1, E. Czekajska-Chehab6, A. Drop6 1Department of Human Anatomy, Medical University of Lublin, Lublin, Poland 2St. John’s Cancer Centre, Lublin, Poland 3Department of Clinical Pathomorphology, Medical University of Lublin, Lublin, Poland 4Department of Anatomy, Collegium Medicum, Jagiellonian University, Krakow, Poland 5Department of Psychiatry and Behavioural Sciences, Behavioural Health Centre, New York Medical College, Valhalla NY, USA 6Department of General Radiology and Nuclear Medicine, Medical University of Lublin, Lublin, Poland [Received 21 July 2012; Accepted 7 September 2012] Background: The foramen magnum is an important anatomical opening in the base of the skull through which the posterior cranial fossa communicates with the vertebral canal. It is also related to a number of pathological condi- tions including Chiari malformations, various tumours, and occipital dysplasias. The aim of the study was to evaluate the morphology of the foramen magnum in adult individuals in relation to sex. Material and methods: The morphology of the foramen magnum was evalu- ated using 3D computer tomography images in 313 individuals (142 male, 171 female) aged 20–30 years. Results: The mean values of the foramen length (37.06 ± 3.07 vs. 35.47 ± ± 2.60 mm), breadth (32.98 ± 2.78 vs. 30.95 ± 2.71 mm) and area (877.40 ± ± 131.64 vs. -

Required List of Bones and Markings
REQUIRED LIST OF BONES AND MARKINGS Axial Skeleton Skull Cranial Bones (8) Frontal Bone (1) Supraorbital foramina Supraorbital ridges or margins Parietal Bones (2) Temporal Bones (2) External auditory meatus Mastoid process Styloid process Zygomatic process Mandibular fossa Foramen lacerum Carotid foramen Jugular foramen Stylomastoid foramen Internal auditory meatus Occipital Bone (1) Foramen magnum Occipital condyles Ethmoid Bone (1) Cribriform plate Olfactory foramina in cribriform plate Crista galli Perpendicular plate (forms superior part of nasal septum) Middle nasal concha Superior nasal concha Sphenoid Bone (1) Foramen ovale Foramen rotundum Sella turcica Greater wing Lesser wing Optic foramen Inferior orbital fissure Superior orbital fissure Pterygoid processes Skull (cont’d) Facial Bones (14) Lacrimal Bones (2) Lacrimal fossa Nasal Bones (2) Inferior Nasal Conchae (2) Vomer (1) (forms inferior portion of nasal septum) Zygomatic Bones (2) Temporal process (forms zygomatic arch with zygomatic process of temporal bone) Maxillae (2) Alveoli Palatine process (forms anterior part of hard palate) Palatine Bones (2) (form posterior part of hard palate) Mandible (1) Alveoli Body Mental foramen Ramus Condylar process (mandibular condyle) Coronoid process Miscellaneous (Skull) Paranasal sinuses are located in the ethmoid bone, sphenoid bone, frontal bone, and maxillae Zygomatic arch (“cheekbone”) is composed of the zygomatic process of the temporal bone and the temporal process of the zygomatic bone 2 pairs of nasal conchae (superior and middle) are part of the ethmoid bone. 1 pair (inferior) are separate facial bones. All the scroll-like conchae project into the lateral walls of the nasal cavity. Hard palate (“roof of mouth”) is composed of 2 palatine processes of the maxillae and the 2 palatine bones (total of 4 fused bones). -
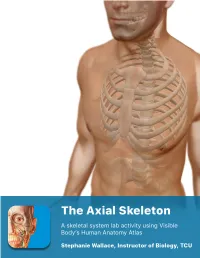
Lab Manual Axial Skeleton Atla
1 PRE-LAB EXERCISES When studying the skeletal system, the bones are often sorted into two broad categories: the axial skeleton and the appendicular skeleton. This lab focuses on the axial skeleton, which consists of the bones that form the axis of the body. The axial skeleton includes bones in the skull, vertebrae, and thoracic cage, as well as the auditory ossicles and hyoid bone. In addition to learning about all the bones of the axial skeleton, it is also important to identify some significant bone markings. Bone markings can have many shapes, including holes, round or sharp projections, and shallow or deep valleys, among others. These markings on the bones serve many purposes, including forming attachments to other bones or muscles and allowing passage of a blood vessel or nerve. It is helpful to understand the meanings of some of the more common bone marking terms. Before we get started, look up the definitions of these common bone marking terms: Canal: Condyle: Facet: Fissure: Foramen: (see Module 10.18 Foramina of Skull) Fossa: Margin: Process: Throughout this exercise, you will notice bold terms. This is meant to focus your attention on these important words. Make sure you pay attention to any bold words and know how to explain their definitions and/or where they are located. Use the following modules to guide your exploration of the axial skeleton. As you explore these bones in Visible Body’s app, also locate the bones and bone markings on any available charts, models, or specimens. You may also find it helpful to palpate bones on yourself or make drawings of the bones with the bone markings labeled. -

MORPHOMETRIC and MORPHOLOGICAL ANALYSIS of FORAMEN OVALE in DRY HUMAN SKULLS Ashwini
International Journal of Anatomy and Research, Int J Anat Res 2017, Vol 5(1):3547-51. ISSN 2321-4287 Original Research Article DOI: https://dx.doi.org/10.16965/ijar.2017.109 MORPHOMETRIC AND MORPHOLOGICAL ANALYSIS OF FORAMEN OVALE IN DRY HUMAN SKULLS Ashwini. N.S *1, Venkateshu. K.V 2. *1 Assistant Professor, Department Of Anatomy,Sri Devaraj Urs Medical College ,Tamaka, Kolar, Karnataka, India. 2 Professor And Head, Department Of Anatomy ,Sri Devaraj Urs Medical College, Tamaka, Kolar, India. ABSTRACT Introduction: Foramen ovale is an important foramen in the middle cranial fossa. Foramen ovale is situated in the greater wing of sphenoid bone, posterior to the foramen rotandum. Through the foramen ovale the mandibular nerve, accessory meningeal artery, lesser petrosal nerve, emissary veins pass .The shape of the foramen ovale is usually oval compared to other foramen of the skull. Materials and Methods: The study was conducted on 55 dry human skulls(110 sides) in Department of Anatomy, Sri Devaraj Urs Medical college, Tamaka, Kolar. Skulls in poor condition or skulls with partially damaged surroundings around the foramen ovale were excluded from the study. Linear measurements were taken on right and left sides of foramen ovale using divider and meter rule. Results: The maximum length of foramen ovale was 14 mm on the right side and 17 mm on the left, its maximum breadth on the right side was 8mm and 10mm on the left . Through the statistical analysis of morphometric measurements between right and left foramen ovale which was found to be insignificant , the results of both sides marks as the evidence of asymmetry in the morphometry of the foramen ovale. -

Microvertebrates of the Lourinhã Formation (Late Jurassic, Portugal)
Alexandre Renaud Daniel Guillaume Licenciatura em Biologia celular Mestrado em Sistemática, Evolução, e Paleobiodiversidade Microvertebrates of the Lourinhã Formation (Late Jurassic, Portugal) Dissertação para obtenção do Grau de Mestre em Paleontologia Orientador: Miguel Moreno-Azanza, Faculdade de Ciências e Tecnologia da Universidade Nova de Lisboa Co-orientador: Octávio Mateus, Faculdade de Ciências e Tecnologia da Universidade Nova de Lisboa Júri: Presidente: Prof. Doutor Paulo Alexandre Rodrigues Roque Legoinha (FCT-UNL) Arguente: Doutor Hughes-Alexandres Blain (IPHES) Vogal: Doutor Miguel Moreno-Azanza (FCT-UNL) Júri: Dezembro 2018 MICROVERTEBRATES OF THE LOURINHÃ FORMATION (LATE JURASSIC, PORTUGAL) © Alexandre Renaud Daniel Guillaume, FCT/UNL e UNL A Faculdade de Ciências e Tecnologia e a Universidade Nova de Lisboa tem o direito, perpétuo e sem limites geográficos, de arquivar e publicar esta dissertação através de exemplares impressos reproduzidos em papel ou de forma digital, ou por qualquer outro meio conhecido ou que venha a ser inventado, e de a divulgar através de repositórios científicos e de admitir a sua cópia e distribuição com objetivos educacionais ou de investigação, não comerciais, desde que seja dado crédito ao autor e editor. ACKNOWLEDGMENTS First of all, I would like to dedicate this thesis to my late grandfather “Papi Joël”, who wanted to tie me to a tree when I first start my journey to paleontology six years ago, in Paris. And yet, he never failed to support me at any cost, even if he did not always understand what I was doing and why I was doing it. He is always in my mind. Merci papi ! This master thesis has been one-year long project during which one there were highs and lows. -

Surgical Management of Posterior Petrous Meningiomas
Neurosurg Focus 14 (6):Article 7, 2003, Click here to return to Table of Contents Surgical management of posterior petrous meningiomas JAMES K. LIU, M.D., OREN N. GOTTFRIED, M.D., AND WILLIAM T. COULDWELL, M.D., PH.D. Department of Neurosurgery, University of Utah School of Medicine, Salt Lake City, Utah Posterior petrous meningiomas (commonly termed posterior pyramid meningiomas and/or meningiomas of the pos- terior surface of the petrous pyramid) are the most common meningiomas of the posterior cranial fossa. They are locat- ed along the posterior surface of the temporal bone in the region of the cerebellopontine angle. They often mimic vestibular schwannomas, both clinically and on neuroimaging studies. Common clinical symptoms include hearing loss, cerebellar ataxia, and trigeminal neuropathy. The site of dural origin determines the direction of cranial nerve dis- placement. Total resection can be achieved in most cases with a low morbidity rate and an excellent prognosis. The authors review the surgical management of posterior petrous meningiomas. KEY WORDS • meningioma • cerebellopontine angle • skull base surgery • petrous bone Posterior fossa meningiomas comprise approximately In their series Samii and Ammirati18 used the term “pos- 10% of all intracranial meningiomas.8 Castellano and terior pyramid meningiomas” and defined them as tumors Ruggiero4 reviewed Olivecrona’s experience with treat- whose main direction of growth brings them in contact ing posterior fossa meningiomas and classified them with the posterior pyramid, irrespective of their site of based on the site of dural attachment. They described the dural attachment. They also designated those located ante- location as cerebellar convexity (10%), tentorium (30%), rior and those posterior to the internal acoustic meatus. -

Topographical Anatomy and Morphometry of the Temporal Bone of the Macaque
Folia Morphol. Vol. 68, No. 1, pp. 13–22 Copyright © 2009 Via Medica O R I G I N A L A R T I C L E ISSN 0015–5659 www.fm.viamedica.pl Topographical anatomy and morphometry of the temporal bone of the macaque J. Wysocki 1Clinic of Otolaryngology and Rehabilitation, II Medical Faculty, Warsaw Medical University, Poland, Kajetany, Nadarzyn, Poland 2Laboratory of Clinical Anatomy of the Head and Neck, Institute of Physiology and Pathology of Hearing, Poland, Kajetany, Nadarzyn, Poland [Received 7 July 2008; Accepted 10 October 2008] Based on the dissections of 24 bones of 12 macaques (Macaca mulatta), a systematic anatomical description was made and measurements of the cho- sen size parameters of the temporal bone as well as the skull were taken. Although there is a small mastoid process, the general arrangement of the macaque’s temporal bone structures is very close to that which is observed in humans. The main differences are a different model of pneumatisation and the presence of subarcuate fossa, which possesses considerable dimensions. The main air space in the middle ear is the mesotympanum, but there are also additional air cells: the epitympanic recess containing the head of malleus and body of incus, the mastoid cavity, and several air spaces on the floor of the tympanic cavity. The vicinity of the carotid canal is also very well pneuma- tised and the walls of the canal are very thin. The semicircular canals are relatively small, very regular in shape, and characterized by almost the same dimensions. The bony walls of the labyrinth are relatively thin.