Parasites of Jellyfish in Eastern Australia
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Appendix to Taxonomic Revision of Leopold and Rudolf Blaschkas' Glass Models of Invertebrates 1888 Catalogue, with Correction
http://www.natsca.org Journal of Natural Science Collections Title: Appendix to Taxonomic revision of Leopold and Rudolf Blaschkas’ Glass Models of Invertebrates 1888 Catalogue, with correction of authorities Author(s): Callaghan, E., Egger, B., Doyle, H., & E. G. Reynaud Source: Callaghan, E., Egger, B., Doyle, H., & E. G. Reynaud. (2020). Appendix to Taxonomic revision of Leopold and Rudolf Blaschkas’ Glass Models of Invertebrates 1888 Catalogue, with correction of authorities. Journal of Natural Science Collections, Volume 7, . URL: http://www.natsca.org/article/2587 NatSCA supports open access publication as part of its mission is to promote and support natural science collections. NatSCA uses the Creative Commons Attribution License (CCAL) http://creativecommons.org/licenses/by/2.5/ for all works we publish. Under CCAL authors retain ownership of the copyright for their article, but authors allow anyone to download, reuse, reprint, modify, distribute, and/or copy articles in NatSCA publications, so long as the original authors and source are cited. TABLE 3 – Callaghan et al. WARD AUTHORITY TAXONOMY ORIGINAL SPECIES NAME REVISED SPECIES NAME REVISED AUTHORITY N° (Ward Catalogue 1888) Coelenterata Anthozoa Alcyonaria 1 Alcyonium digitatum Linnaeus, 1758 2 Alcyonium palmatum Pallas, 1766 3 Alcyonium stellatum Milne-Edwards [?] Sarcophyton stellatum Kükenthal, 1910 4 Anthelia glauca Savigny Lamarck, 1816 5 Corallium rubrum Lamarck Linnaeus, 1758 6 Gorgonia verrucosa Pallas, 1766 [?] Eunicella verrucosa 7 Kophobelemon (Umbellularia) stelliferum -

High Level Environmental Screening Study for Offshore Wind Farm Developments – Marine Habitats and Species Project
High Level Environmental Screening Study for Offshore Wind Farm Developments – Marine Habitats and Species Project AEA Technology, Environment Contract: W/35/00632/00/00 For: The Department of Trade and Industry New & Renewable Energy Programme Report issued 30 August 2002 (Version with minor corrections 16 September 2002) Keith Hiscock, Harvey Tyler-Walters and Hugh Jones Reference: Hiscock, K., Tyler-Walters, H. & Jones, H. 2002. High Level Environmental Screening Study for Offshore Wind Farm Developments – Marine Habitats and Species Project. Report from the Marine Biological Association to The Department of Trade and Industry New & Renewable Energy Programme. (AEA Technology, Environment Contract: W/35/00632/00/00.) Correspondence: Dr. K. Hiscock, The Laboratory, Citadel Hill, Plymouth, PL1 2PB. [email protected] High level environmental screening study for offshore wind farm developments – marine habitats and species ii High level environmental screening study for offshore wind farm developments – marine habitats and species Title: High Level Environmental Screening Study for Offshore Wind Farm Developments – Marine Habitats and Species Project. Contract Report: W/35/00632/00/00. Client: Department of Trade and Industry (New & Renewable Energy Programme) Contract management: AEA Technology, Environment. Date of contract issue: 22/07/2002 Level of report issue: Final Confidentiality: Distribution at discretion of DTI before Consultation report published then no restriction. Distribution: Two copies and electronic file to DTI (Mr S. Payne, Offshore Renewables Planning). One copy to MBA library. Prepared by: Dr. K. Hiscock, Dr. H. Tyler-Walters & Hugh Jones Authorization: Project Director: Dr. Keith Hiscock Date: Signature: MBA Director: Prof. S. Hawkins Date: Signature: This report can be referred to as follows: Hiscock, K., Tyler-Walters, H. -

THE J!IEDUS..E of the WOODS HOLE REGION
CONTRIBl"T10NS FROM TII£ BIOLOGICAL LABORATORY or THE BUREAU OF FISIIERIES AT WOODS HOLE, MASS. THE j!IEDUS ..E OF THE WOODS HOLE REGION. 13y CI1ANLES vv, I-IA H CHTT, Prokssor of7oo1oK,J'. Svracuse {/lli7'ersity. 21 CONTRIBUTIONS FRO&{ THE BIOLOGICAl. LABORATORY OF TilE HUREAL OF FlsHElm:S AT WOODS HOLE, MASSACHUSliTI'S, THE jIEDUS.E 01; THE WOO])S HOLE REGION. By CHARLES W. HARGITT, ProkHor (~/ X,)O!O(;y, ,))'1'<101.1'" Cni"a:,itv. INTRODUCTION. The present report f'onns one of' a ser-ies projected hy the director of the biological laboratory of the United States Bun~au of' Fbheric:--, the prirnarv oh,it'd lwill!! to aflord such a biological survey of the region as will bring' within l~a"y I'('lwh of students and working naturalist" a synopsis of tlw eharaeter and distribution of its fauna. The work which forms the basis of this pap.,!, was cluTied on during- t/;p summers of'HHII and l!102, including' also a brief ('olleeting recllllllais"alH'C during the early ,"'pring of' the latter year, thus l'nabling me to eomp!l'b' :l recor-d (If observations upon the medusoid fauna during' pvcry month of t.ho yl'i:lJ', with daily record" dUJ'ing most of the time. FOI' part" of th(',,(' l'l'l'ol'd,s during' lut.- fall aud winterI all! chiefly indebted to 1\11'. Viuul X. Edwanl", which it i,; a ph'aKlIl'p hl'I'P!ly to acknowledge, It is also a pleasure toacknowll'dge tlH' eordiaI1'oolH'l'atioll of' the Commissioner, Hon. -

Anthopleura and the Phylogeny of Actinioidea (Cnidaria: Anthozoa: Actiniaria)
Org Divers Evol (2017) 17:545–564 DOI 10.1007/s13127-017-0326-6 ORIGINAL ARTICLE Anthopleura and the phylogeny of Actinioidea (Cnidaria: Anthozoa: Actiniaria) M. Daly1 & L. M. Crowley2 & P. Larson1 & E. Rodríguez2 & E. Heestand Saucier1,3 & D. G. Fautin4 Received: 29 November 2016 /Accepted: 2 March 2017 /Published online: 27 April 2017 # Gesellschaft für Biologische Systematik 2017 Abstract Members of the sea anemone genus Anthopleura by the discovery that acrorhagi and verrucae are are familiar constituents of rocky intertidal communities. pleisiomorphic for the subset of Actinioidea studied. Despite its familiarity and the number of studies that use its members to understand ecological or biological phe- Keywords Anthopleura . Actinioidea . Cnidaria . Verrucae . nomena, the diversity and phylogeny of this group are poor- Acrorhagi . Pseudoacrorhagi . Atomized coding ly understood. Many of the taxonomic and phylogenetic problems stem from problems with the documentation and interpretation of acrorhagi and verrucae, the two features Anthopleura Duchassaing de Fonbressin and Michelotti, 1860 that are used to recognize members of Anthopleura.These (Cnidaria: Anthozoa: Actiniaria: Actiniidae) is one of the most anatomical features have a broad distribution within the familiar and well-known genera of sea anemones. Its members superfamily Actinioidea, and their occurrence and exclu- are found in both temperate and tropical rocky intertidal hab- sivity are not clear. We use DNA sequences from the nu- itats and are abundant and species-rich when present (e.g., cleus and mitochondrion and cladistic analysis of verrucae Stephenson 1935; Stephenson and Stephenson 1972; and acrorhagi to test the monophyly of Anthopleura and to England 1992; Pearse and Francis 2000). -

Luth Wfu 0248D 10922.Pdf
SCALE-DEPENDENT VARIATION IN MOLECULAR AND ECOLOGICAL PATTERNS OF INFECTION FOR ENDOHELMINTHS FROM CENTRARCHID FISHES BY KYLE E. LUTH A Dissertation Submitted to the Graduate Faculty of WAKE FOREST UNIVERSITY GRADAUTE SCHOOL OF ARTS AND SCIENCES in Partial Fulfillment of the Requirements for the Degree of DOCTOR OF PHILOSOPHY Biology May 2016 Winston-Salem, North Carolina Approved By: Gerald W. Esch, Ph.D., Advisor Michael V. K. Sukhdeo, Ph.D., Chair T. Michael Anderson, Ph.D. Herman E. Eure, Ph.D. Erik C. Johnson, Ph.D. Clifford W. Zeyl, Ph.D. ACKNOWLEDGEMENTS First and foremost, I would like to thank my PI, Dr. Gerald Esch, for all of the insight, all of the discussions, all of the critiques (not criticisms) of my works, and for the rides to campus when the North Carolina weather decided to drop rain on my stubborn head. The numerous lively debates, exchanges of ideas, voicing of opinions (whether solicited or not), and unerring support, even in the face of my somewhat atypical balance of service work and dissertation work, will not soon be forgotten. I would also like to acknowledge and thank the former Master, and now Doctor, Michael Zimmermann; friend, lab mate, and collecting trip shotgun rider extraordinaire. Although his need of SPF 100 sunscreen often put our collecting trips over budget, I could not have asked for a more enjoyable, easy-going, and hard-working person to spend nearly 2 months and 25,000 miles of fishing filled days and raccoon, gnat, and entrail-filled nights. You are a welcome camping guest any time, especially if you do as good of a job attracting scorpions and ants to yourself (and away from me) as you did on our trips. -

1 Curriculum Vitae Stephen S. Curran, Ph.D. Department of Coastal
Curriculum vitae Stephen S. Curran, Ph.D. Department of Coastal Sciences The University of Southern Mississippi Gulf Coast Research Laboratory 703 East Beach Drive Phone: (228) 238-0208 Ocean Springs, MS 39564 Email: [email protected] Research and Teaching Interests: I am an organismal biologist interested in the biodiversity of metazoan parasitic animals. I study their taxonomy using traditional microscopic and histological techniques and their genetic interrelationships and systematics using ribosomal DNA sequences. I also investigate the effects of extrinsic factors on aquatic environments by using parasite prevalence and abundance as a proxy for total biodiversity in aquatic communities and for assessing food web dynamics. I am also interested in the epidemiology of viral diseases of crustaceans. University Teaching Experience: •Instructor for Parasites of Marine Animals Summer class, University of Southern Mississippi, Gulf Coast Research Laboratory (2011-present). •Co-Instructor (with Richard Heard) for Marine Invertebrate Zoology, University of Southern Mississippi, Gulf Coast Research Laboratory (2007). •Intern Mentor, Gulf Coast Research Laboratory. I’ve instructed 16 interns during (2003, 2007- present). •Graduate Teaching Assistant for Animal Parasitology, Department of Ecology and Evolutionary Biology, University of Connecticut (Spring 1995). •Graduate Teaching Assistant for Introductory Biology for Majors, Department of Ecology and Evolutionary Biology, University of Connecticut (Fall 1994). Positions: •Assistant Research -

Digenean Metacercaria (Trematoda, Digenea, Lepocreadiidae) Parasitizing “Coelenterates” (Cnidaria, Scyphozoa and Ctenophora) from Southeastern Brazil
BRAZILIAN JOURNAL OF OCEANOGRAPHY, 53(1/2):39-45, 2005 DIGENEAN METACERCARIA (TREMATODA, DIGENEA, LEPOCREADIIDAE) PARASITIZING “COELENTERATES” (CNIDARIA, SCYPHOZOA AND CTENOPHORA) FROM SOUTHEASTERN BRAZIL André Carrara Morandini1; Sergio Roberto Martorelli2; Antonio Carlos Marques1 & Fábio Lang da Silveira1 1Instituto de Biociências da Universidade de São Paulo Departamento de Zoologia (Caixa Postal 11461, 05422-970, São Paulo, SP, Brazil) 2Centro de Estudios Parasitologicos y Vectores (CEPAVE) (2 Nro. 584 (1900) La Plata, Argentina) e-mails: [email protected], [email protected], [email protected], [email protected] A B S T R A C T Metacercaria specimens of the genus Opechona (Trematoda: Digenea: Lepocreadiidae) are described parasitizing “coelenterates” (scyphomedusae and ctenophores) from Southeastern Brazil (São Paulo state). The worms are compared to other Opechona species occurring on the Brazilian coast, but no association has been made because only adult forms of these species have been described. Suppositions as to the possible transference of the parasites are made. R E S U M O Exemplares de metacercárias do gênero Opechona (Trematoda: Digenea: Lepocreadiidae) são descritos parasitando “celenterados” (cifomedusas e ctenóforos) no sudeste do Brasil (estado de São Paulo). Os vermes foram comparados a outras espécies de Opechona ocorrentes no litoral brasileiro, porém nenhuma associação foi realizada devido às demais espécies terem sido descritas a partir de exemplares adultos. São apresentadas suposições sobre as possíveis formas -

Parasites of Coral Reef Fish: How Much Do We Know? with a Bibliography of Fish Parasites in New Caledonia
Belg. J. Zool., 140 (Suppl.): 155-190 July 2010 Parasites of coral reef fish: how much do we know? With a bibliography of fish parasites in New Caledonia Jean-Lou Justine (1) UMR 7138 Systématique, Adaptation, Évolution, Muséum National d’Histoire Naturelle, 57, rue Cuvier, F-75321 Paris Cedex 05, France (2) Aquarium des lagons, B.P. 8185, 98807 Nouméa, Nouvelle-Calédonie Corresponding author: Jean-Lou Justine; e-mail: [email protected] ABSTRACT. A compilation of 107 references dealing with fish parasites in New Caledonia permitted the production of a parasite-host list and a host-parasite list. The lists include Turbellaria, Monopisthocotylea, Polyopisthocotylea, Digenea, Cestoda, Nematoda, Copepoda, Isopoda, Acanthocephala and Hirudinea, with 580 host-parasite combinations, corresponding with more than 370 species of parasites. Protozoa are not included. Platyhelminthes are the major group, with 239 species, including 98 monopisthocotylean monogeneans and 105 digeneans. Copepods include 61 records, and nematodes include 41 records. The list of fish recorded with parasites includes 195 species, in which most (ca. 170 species) are coral reef associated, the rest being a few deep-sea, pelagic or freshwater fishes. The serranids, lethrinids and lutjanids are the most commonly represented fish families. Although a list of published records does not provide a reliable estimate of biodiversity because of the important bias in publications being mainly in the domain of interest of the authors, it provides a basis to compare parasite biodiversity with other localities, and especially with other coral reefs. The present list is probably the most complete published account of parasite biodiversity of coral reef fishes. -
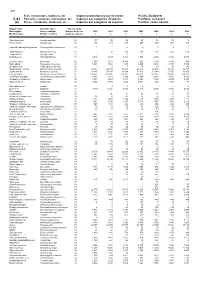
Fish, Crustaceans, Molluscs, Etc Capture Production by Species
478 Fish, crustaceans, molluscs, etc Capture production by species items Pacific, Southwest C-81 Poissons, crustacés, mollusques, etc Captures par catégories d'espèces Pacifique, sud-ouest (a) Peces, crustáceos, moluscos, etc Capturas por categorías de especies Pacífico, sudoccidental English name Scientific name Species group Nom anglais Nom scientifique Groupe d'espèces 2000 2001 2002 2003 2004 2005 2006 Nombre inglés Nombre científico Grupo de especies t t t t t t t Short-finned eel Anguilla australis 22 37 38 28 27 13 10 5 River eels nei Anguilla spp 22 380 313 337 267 209 277 210 Chinook(=Spring=King)salmon Oncorhynchus tshawytscha 23 1 1 0 4 1 2 1 Sand flounders Rhombosolea spp 31 ... 37 204 193 187 437 514 Tonguefishes Cynoglossidae 31 - - 3 - - - - Flatfishes nei Pleuronectiformes 31 2 954 3 234 2 818 3 308 2 980 3 766 3 050 Common mora Mora moro 32 1 358 1 211 1 308 1 234 1 403 1 154 986 Red codling Pseudophycis bachus 32 5 364 4 526 4 443 8 265 9 540 8 165 5 854 Grenadier cod Tripterophycis gilchristi 32 - 2 7 10 13 13 43 Southern blue whiting Micromesistius australis 32 43 419 49 982 72 203 43 812 26 576 30 304 32 735 Southern hake Merluccius australis 32 16 041 15 188 13 834 22 623 19 344 12 560 12 858 Blue grenadier Macruronus novaezelandiae 32 274 625 247 841 215 302 209 414 147 032 134 145 119 329 Thorntooth grenadier Lepidorhynchus denticulatus 32 3 833 4 783 5 349 5 304 6 341 3 855 4 056 Grenadiers, rattails nei Macrouridae 32 2 394 3 094 3 877 4 253 3 732 2 660 2 848 Gadiformes nei Gadiformes 32 2 853 5 479 3 252 3 281 298 1 217 47 Sea catfishes nei Ariidae 33 .. -

Checklist of Marine Demersal Fishes Captured by the Pair Trawl Fisheries in Southern (RJ-SC) Brazil
Biota Neotropica 19(1): e20170432, 2019 www.scielo.br/bn ISSN 1676-0611 (online edition) Inventory Checklist of marine demersal fishes captured by the pair trawl fisheries in Southern (RJ-SC) Brazil Matheus Marcos Rotundo1,2,3,4 , Evandro Severino-Rodrigues2, Walter Barrella4,5, Miguel Petrere Jun- ior3 & Milena Ramires4,5 1Universidade Santa Cecilia, Acervo Zoológico, R. Oswaldo Cruz, 266, CEP11045-907, Santos, SP, Brasil 2Instituto de Pesca, Programa de Pós-graduação em Aquicultura e Pesca, Santos, SP, Brasil 3Universidade Federal de São Carlos, Programa de Pós-Graduação em Planejamento e Uso de Recursos Renováveis, Rodovia João Leme dos Santos, Km 110, CEP 18052-780, Sorocaba, SP, Brasil 4Universidade Santa Cecília, Programa de Pós-Graduação de Auditoria Ambiental, R. Oswaldo Cruz, 266, CEP11045-907, Santos, SP, Brasil 5Universidade Santa Cecília, Programa de Pós-Graduação em Sustentabilidade de Ecossistemas Costeiros e Marinhos, R. Oswaldo Cruz, 266, CEP11045-907, Santos, SP, Brasil *Corresponding author: Matheus Marcos Rotundo: [email protected] ROTUNDO, M.M., SEVERINO-RODRIGUES, E., BARRELLA, W., PETRERE JUNIOR, M., RAMIRES, M. Checklist of marine demersal fishes captured by the pair trawl fisheries in Southern (RJ-SC) Brazil. Biota Neotropica. 19(1): e20170432. http://dx.doi.org/10.1590/1676-0611-BN-2017-0432 Abstract: Demersal fishery resources are abundant on continental shelves, on the tropical and subtropical coasts, making up a significant part of the marine environment. Marine demersal fishery resources are captured by various fishing methods, often unsustainably, which has led to the depletion of their stocks. In order to inventory the marine demersal ichthyofauna on the Southern Brazilian coast, as well as their conservation status and distribution, this study analyzed the composition and frequency of occurrence of fish captured by pair trawling in 117 fishery fleet landings based in the State of São Paulo between 2005 and 2012. -

The Trematode Parasites of Marine Mammals
THE TREMATODE PARASITES OF MARINE MAMMALS By Emmett W. Pkice Parasitologist, Zoological Division, Bureau of Animal Industry United States Department of Agriculture The internal parasites of marine mammals have not been exten- sively studied, although a fairly large number of species have been described. In attempting to identify the trematodes from mammals of the orders Cetacea, Pinnipedia, and Sirenia, as represented by specimens in the United States National Museum helminthological collection, it was necessary to review the greater part of the litera- ture dealing with this group of parasitic worms. In view of the fact that there is not in existence a single comprehensive paper on the trematodes of these mammals, and that many of the descrip- tions of species have appeared in publications having more or less limited circulation, the writer has undertaken to assemble descriptions of all trematodes reported from these hosts, with the hope that such a paper may serve a useful purpose in aiding other workers in de- termining specimens at their disposal. In addition to compiling the descriptions of species not available to the writer, two new species, one of which represents a new genus, have been described. Specimens representing 10 of the previously described species have been studied and emendations or additions have been made to the existing descriptions; in a few instances the species have been completely reclescribed. Three species, Distoinwni pallassil Poirier, D. vaUdwim von Lin- stow, and D. am/pidlacewni Buttel-Reepen, have been omitted from this paper despite the fact that they have been reported from ceta- ceans. These species belong in the family Hemiuridae, and since all species of this family are parasites of fishes, the writer feels that their reported occurrence in mammals may be regarded as either errors of some sort or cases of accidental parasitism in which fishes have been eaten by mammals and the fish parasites found in the mammal post-mortem. -

Platyhelminthes: Tricladida: Terricola) of the Australian Region
ResearchOnline@JCU This file is part of the following reference: Winsor, Leigh (2003) Studies on the systematics and biogeography of terrestrial flatworms (Platyhelminthes: Tricladida: Terricola) of the Australian region. PhD thesis, James Cook University. Access to this file is available from: http://eprints.jcu.edu.au/24134/ The author has certified to JCU that they have made a reasonable effort to gain permission and acknowledge the owner of any third party copyright material included in this document. If you believe that this is not the case, please contact [email protected] and quote http://eprints.jcu.edu.au/24134/ Studies on the Systematics and Biogeography of Terrestrial Flatworms (Platyhelminthes: Tricladida: Terricola) of the Australian Region. Thesis submitted by LEIGH WINSOR MSc JCU, Dip.MLT, FAIMS, MSIA in March 2003 for the degree of Doctor of Philosophy in the Discipline of Zoology and Tropical Ecology within the School of Tropical Biology at James Cook University Frontispiece Platydemus manokwari Beauchamp, 1962 (Rhynchodemidae: Rhynchodeminae), 40 mm long, urban habitat, Townsville, north Queensland dry tropics, Australia. A molluscivorous species originally from Papua New Guinea which has been introduced to several countries in the Pacific region. Common. (photo L. Winsor). Bipalium kewense Moseley,1878 (Bipaliidae), 140mm long, Lissner Park, Charters Towers, north Queensland dry tropics, Australia. A cosmopolitan vermivorous species originally from Vietnam. Common. (photo L. Winsor). Fletchamia quinquelineata (Fletcher & Hamilton, 1888) (Geoplanidae: Caenoplaninae), 60 mm long, dry Ironbark forest, Maryborough, Victoria. Common. (photo L. Winsor). Tasmanoplana tasmaniana (Darwin, 1844) (Geoplanidae: Caenoplaninae), 35 mm long, tall open sclerophyll forest, Kamona, north eastern Tasmania, Australia.