The Effect of Plant Hormones on Phenolic Production in Kauri Trees
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-
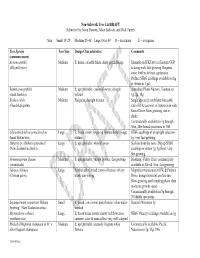
Recommended Non-Sidewalk Tree List DRAFT
Non-Sidewalk Tree List DRAFT Submitted by Jason Dewees, Mike Sullivan, and Dick Turner Size: Small 15-25’ Medium 25-40’ Large Over 40’ D = deciduous E = evergreen Tree Species Tree Size Design Characteristics Comments (common name) Acmena smithii Medium E; dense, colorful fruits; shiny green foliage Example on JFK Drive in Eastern GGP (lilly-pilly tree) is doing well; fast-growing. Requires some fertility for best appearance. Profuse SFBG seedlings available to dig or obtain in 1 gal. Banksia integrifolia Medium E; upright habit; creamy flowers; drought Australian Plants Nursery, Ventura, in (coast banksia) tolerant 1g, 5g, 15g Brahea edulis Medium Fan palm; drought tolerant Single specimen established on north (Guadalupe palm) side of JFK just west of intersection with Kezar Drive. Slow growing, sun or shade. Commercially available in 1g through 36in, 48in boxed specimens to 15ft Chiranthodendron pentadactylon Large E; broad crown; large red flowers, bold foliage SFBG seedlings of an upright selection: (hand flower tree) texture 1g; very fast-growing Hoheria sp. (Hoheria populnea? Large E; upright habit; white flowers Suckers from the roots. Dig up SFBG (New Zealand lacebark)) seedlings or obtain 1g, 5g there; very fast-growing Hymenosporum flavum Medium E; upright habit; yellow flowers; fast growing Boething, Valley Crest; commercially (sweetshade) available in 24in & 36in; fast-growing. Jubaea chilensis Large Feather palm; broad crown of leaves; silvery Magnificent specimen at JFK & Fuchsia (Chilean palm) trunk; sun-loving Drive; drought-tolerant, prefers sun. Slow-growing until trunking phase, then moderate growth speed. Commercially available in 5g through 20ft B&B specimens Leptospermum scoparium ‘Helene Small E; broad, low crown; pink flowers; often multi- Suncrest Nurseries 5g Strybing’ (New Zealand tea tree) trunked Metrosideros robusta Large E; broad dense crown; showy red flowers in SFBG Nursery seedlings available in 1g (northern rata) summer; slow & unusual but very well-adapted Michelia/Magnolia champaca or M. -

Agathis Robusta and Agathis Australis Friends Friends
Plants in Focus, December 2016 Agathis robusta and Agathis australis Friends of GeelongBotanic Left: The Qld Kauri Agathis robusta, planted in the Albury BG in 1910, is the largest recorded in the Big Tree Register. Note gardener. [1] Right: The NZ Kauri Agathis australis, named Tane Mahuta (Lord of the Forest), in the Waipoua Forest is the largest known in NZ. Photo: Prof. Chen Hualin, CC BY-SA 4.0, zh.wikipedia.org Kauris (Agathis sp.) are conifers Conifers, along with the other Gymnosperms (Cycads and Ginkgoes), first appeared about 300 Ma (Million years ago) at the end of the Carboniferous when the world’s coal deposits were being laid down with the remains of the spore-producing trees of that period. The early conifers looked like modern Araucaria. These trees spread throughout the world and displaced their predecessors. The age of the seed plants had arrived. The conifers are a hardy lot. They survived the largest mass extinction the earth has known, 252 Ma, at the end of the Permian Period. But more challenges lay ahead. Sometime in the next 50 Myr (Million years) one of Gymnosperms gave rise to the flowering plants, the Angiosperms. By 100 Ma, in the Cretaceous period, Angiosperms were widespread. And so the battle began - and still continues to this day. The flowering plants have many features that make them more successful in many environments, so their take-over of many habitats was complete by about 65 Ma at the end of the age of the dinosaurs. But in the world’s harsh environments the conifers continue to not just survive, but flourish. -

Phytoalexins: Current and Possible Future Applications in Human Health and Diseases Control
International Journal of Molecular Biology: Open Access Review Article Open Access Phytoalexins: Current and possible future applications in human health and diseases control Abstract Volume 3 Issue 3 - 2018 Plants are prone to diseases and infections following their obvious exposure to microbes Anthony Cemaluk C Egbuonu,1 Juliet C and attendant microbial attacks. Plants control diseases and infections by using their 2 secondary metabolites known collectively as phytoalexins. These phytoalexins, usually Eneogwe 1 synthesized in plants in response to diseases and infections, have enormous chemical Department of Biochemistry, Michael Okpara University of Agriculture, Nigeria diversity and biologic roles but are essentially non biodegradable owing to their stable 2Department of Biochemistry, Michael Okpara University of structures hence could bio-accumulate with sustained effect once synthesized. Thus, Agriculture, Nigeria this reviewed current and possible future applications of phytoalexins in human health and diseases control using relevant search words and search engines. The review noted Correspondence: Anthony Cemaluk C Egbuonu, Department that resveratrol, a representative of, and an extensively studied, phytoalexins was of Biochemistry, Michael Okpara University of Agriculture, variously implicated in the management of human health, including in the prevention Umudike, Abia State, Nigeria, Tel +23480.3636-6565, of cardiovascular disease and cancers. Resveratrol acts via mechanisms essentially Email [email protected] related to its capacity to ameliorate oxidative stress perhaps by significantly enhancing the synthesis of nitric oxide, NO, which could act as an antioxidant. Received: October 30, 2017 | Published: May 10, 2018 Increased oxidative stress has been implicated in human diseases and efforts aimed at mitigating (or preventing the onset of) oxidative stress have been the underlying approach to human disease management and control. -

26 Extreme Trees Pub 2020
Publication WSFNR-20-22C April 2020 Extreme Trees: Tallest, Biggest, Oldest Dr. Kim D. Coder, Professor of Tree Biology & Health Care / University Hill Fellow University of Georgia Warnell School of Forestry & Natural Resources Trees have a long relationship with people. They are both utility and amenity. Trees can evoke awe, mysticism, and reverence. Trees represent great public and private values. Trees most noticed and celebrated by people and communities are the one-tenth of one-percent of trees which approach the limits of their maximum size, reach, extent, and age. These singular, historic, culturally significant, and massive extreme trees become symbols and icons of life on Earth, and our role model in environmental stewardship and sustainability. What Is A Tree? Figure 1 is a conglomeration of definitions and concepts about trees from legal and word definitions in North America. For example, 20 percent of all definitions specifically state a tree is a plant. Concentrated in 63% of all descriptors for trees are four terms: plant, woody, single stem, and tall. If broad stem diameter, branching, and perennial growth habit concepts are added, 87% of all the descriptors are represented. At its most basic level, defining a tree is not species based, but is a structural definition. A tree is represented by a type of plant architecture recognizable by non-technical people. The most basic concepts for defining a tree are — a large, tall, woody, perennial plant with a single, unbranched, erect, self-supporting stem holding an elevated and distinct crown of branches, and which is greater than 10 feet in height and greater than 3 inches in diameter. -

NOTES on AGATHIS AUSTRALIS. by C
NOTES ON AGATHIS AUSTRALIS. By C. T. SANDO. Introduction.—Much has been published about our kauri (Agathis australis) and it is now generally recognised as one of the finest coniferous trees of the world. The New Zealand species is one of a genus of ten—the most important being— A. Palmerstoni ... (Queensland) A. microstachya ... (Queensland) A. robusta ... (Queensland and Philippine Islands) A. vitiensis * ... (Fiji) A. alba ... (Malaya, Sumatra, Java, Celebes, Borneo, and Philippine Islands) A. lanceolata ... (New Caledonia) A. macrophylla ... (Solomon Islands) The value of Agathis australis was first recognised before 1800 by which time a considerable trade in kauri spars had sprung up between Australia and New Zealand. Later, naval-store ships were sent out from England for cargoes and from that time on kauri has increased in value. Up till a short time ago it was used for building and interior furnishing, but now the limited supplies demand a price far too high for these purposes so that its present uses have been confined to carriage building, cabinet-making, and general joinery, vats, tubs, and other special purposes. Wanton waste has so depleted out kauri areas that now, more than ever it is forced upon us, the necessity of making very serious attempts at conservation of the remaining supplies and propagation of this tree for scenic purposes, and, if economic conditions warrant it, for timber production. Geographical, Climatic, and Edaphic Range.—Kauri occurs in the northern parts of the North Island except in the extreme northern peninsula. It is found from Ahipara and Mangonui in the north as far south as the Bay of Plenty on the East Coast and Kawhia Harbour on the west, i.e. -

The Scientific Reserves of Auckland University. Ii. Quantitative Vegetation Studies
TANE 29, 1983 THE SCIENTIFIC RESERVES OF AUCKLAND UNIVERSITY. II. QUANTITATIVE VEGETATION STUDIES by John Ogden Department of Botany, University of Auckland, Private Bag, Auckland SUMMARY Forest vegetation surveys, using both plots and point-centred-quarter (plotless) methods, were undertaken by students in the University reserves at Swanson, Huapai, Oratia and Leigh over the period 1979 to 1982. The two methods gave similar estimates of species composition, total basal area and total density, but both gave wide confidence intervals on the parameters being estimated. The "leading species", in terms of both density and basal area, are defined for each area. The differences between the rankings for these different vegetation parameters are commented upon A regression of age on height of kauri (Agathis australis) seedlings is presented. The relationship between age and trunk diameter is discussed. It is shown that in cores taken from kauri poles in ricker stands over the size range 10 to 40 cm DBH the relationship between age and diameter is very weak. When a wider size range is employed a highly statistically significant relationship is obtained. It is concluded that the highly skewed size class frequency distribution of kauri, characteristic of such stands, reflects a competitive hierarchy. The senile population structure of kanuka (Leptospermum ericoides) in ricker stands confirms their serai role in forest development. The basal area estimates are discussed in the context of other figures for New Zealand forests. It is concluded that although the values are high (average 70 m2 ha 1) they are nevertheless below those commonly found in mature kauri forest. -

Investigation of Rice Diterpenoid Phytoalexin Biosynthesis Benjamin C
Iowa State University Capstones, Theses and Graduate Theses and Dissertations Dissertations 2017 Investigation of rice diterpenoid phytoalexin biosynthesis Benjamin C. Brown Iowa State University Follow this and additional works at: https://lib.dr.iastate.edu/etd Part of the Biochemistry Commons Recommended Citation Brown, Benjamin C., "Investigation of rice diterpenoid phytoalexin biosynthesis" (2017). Graduate Theses and Dissertations. 16322. https://lib.dr.iastate.edu/etd/16322 This Thesis is brought to you for free and open access by the Iowa State University Capstones, Theses and Dissertations at Iowa State University Digital Repository. It has been accepted for inclusion in Graduate Theses and Dissertations by an authorized administrator of Iowa State University Digital Repository. For more information, please contact [email protected]. Investigation of rice diterpenoid phytoalexin biosynthesis by Benjamin C. Brown A thesis submitted to the graduate faculty in partial fulfillment of the requirements for the degree MASTER OF SCIENCE Major: Biochemistry Program of Study Committee: Reuben J. Peters, Major Professor Gustavo MacIntosh Bing Yang The student author and the program of study committee are solely responsible for the content of this thesis. The graduate college will ensure this thesis is globally accessible and will not permit alterations once a degree is conferred. Iowa State University Ames, IA 2018 Copyright © Benjamin C. Brown, 2018. All rights reserved. ii TABLE OF CONTENTS ABSTRACT iv CHAPTER 1. GENERAL INTRODUCTION 1 References 7 Figures 8 CHAPTER 2. MATERIALS AND METHODS 9 Materials 9 CRISPR/Cas9 Vectors 9 Plant Growth 10 Induction and Extraction of Diterpenoids 11 LC-MS/MS Analysis of Diterpenoids 12 References 15 Figures 15 Tables 30 CHAPTER 3. -

Molecular Biology of Disease Resistance in Rice
Physiological and Molecular Plant Pathology (2001) 59, 1±11 doi:10.1006/pmpp.2001.0353, available online at http://www.idealibrary.com on MINI-REVIEW Molecular biology of disease resistance in rice FENGMING SONG1,2 and ROBERT M. GOODMAN1* 1Department of Plant Pathology, University of Wisconsin, 1630 Linden Drive, Madison, WI 53706, U.S.A. and 2Department of Plant Protection, College of Agriculture and Biotechnology, Zhejiang University, Hangzhou, Zhejiang, 310029, P.R. China (Accepted for publication 13 August 2001) Rice is one of the most important staple foods for the understanding the molecular biology of disease resistance increasing world population, especially in Asia. Diseases in rice is a prerequisite. are among the most important limiting factors that aect In recent years, rice has been recognized as a genetic rice production, causing annual yield loss conservatively model for molecular biology research aimed toward estimated at 5 %. More than 70 diseases caused by fungi, understanding mechanisms for growth, development and bacteria, viruses or nematodes have been recorded on rice stress tolerance as well as disease resistance [34]. Rice as a [68], among which rice blast (Magnaporthe grisea), model crop is a fortuitous situation since it is also a crop of bacterial leaf blight (Xanthomonas oryzae pv. oryzae) and world signi®cance. Rice is an attractive model for plant sheath blight (Rhizoctonia solani) are the most serious genetics and genomics because it has a relatively small constraints on high productivity [68]. Resistant cultivars genome. Considerable progress has been made in rice and application of pesticides have been used for disease towards cloning and identi®cation of disease resistance control. -

Plant Disease Control: Understanding the Roles of Toxins and Phytoalexins in Host-Pathogen Interaction
View metadata, citation and similar papers at core.ac.uk brought to you by CORE provided by Pertanika Journal of Scholarly Research Reviews (PJSRR - Universiti Putra Malaysia,... PJSRR (2018) 4(1): 54-66 eISSN: 2462-2028 © Universiti Putra Malaysia Press Pertanika Journal of Scholarly Research Reviews http://www.pjsrr.upm.edu.my/ Plant Disease Control: Understanding the Roles of Toxins and Phytoalexins in Host-Pathogen Interaction Magaji G USMANa,b, Tijjani AHMADUb, ADAMU Jibrin Nayayab, AISHA M Dodoc aLaboratory of Climate-Smart Food Crop Production, Institute of Tropical Agriculture and Food Security, Universiti Putra Malaysia, 43400 UPM Serdang, Selangor, Malaysia bDepartment of Crop Production, Faculty of Agriculture and Agricultural Technology, Abubakar Tafawa Balewa University (ATBU). PMB 0248, Bauchi, Nigeria cCardiff School of Sport and Health Sciences, Cardiff Metropolitan University, Cardiff, United Kingdom *[email protected] Abstract – Naturally, plant habitats are exposed to several potential effects of biotic and different abiotic environmental challenges. Several types of micro-organisms namely; bacteria, viruses, fungi, nematodes, mites, insects, mammals and other herbivorous animals are found in large amounts in all ecosystems, which lead to considerable reduction in crop productivity. These organisms are agents carrying different diseases that can damage the plants through the secretion of toxic-microbial poisons that can penetrate in the plant tissues. Toxins are injurious substances that act on plant protoplast to influence disease development. In response to the stress effect, plants defend themselves by bearing some substances such as phytoalexins. Production of phytoalexins is one of the complex mechanisms through which plants exhibit disease resistance. Several findings specifically on phytoalexins have widen the understanding in the fields of plant biochemistry and molecular biology. -

Phenolic Compounds in Trees and Shrubs of Central Europe
applied sciences Review Phenolic Compounds in Trees and Shrubs of Central Europe Lidia Szwajkowska-Michałek 1,*, Anna Przybylska-Balcerek 1 , Tomasz Rogozi ´nski 2 and Kinga Stuper-Szablewska 1 1 Department of Chemistry, Faculty of Forestry and Wood Technology, Pozna´nUniversity of Life Sciences ul. Wojska Polskiego 75, 60-625 Pozna´n,Poland; [email protected] (A.P.-B.); [email protected] (K.S.-S.) 2 Department of Furniture Design, Faculty of Forestry and Wood Technology, Pozna´nUniversity of Life Sciences ul. Wojska Polskiego 38/42, 60-627 Pozna´n,Poland; [email protected] * Correspondence: [email protected]; Tel.: +48-61-848-78-43 Received: 1 September 2020; Accepted: 30 September 2020; Published: 2 October 2020 Abstract: Plants produce specific structures constituting barriers, hindering the penetration of pathogens, while they also produce substances inhibiting pathogen growth. These compounds are secondary metabolites, such as phenolics, terpenoids, sesquiterpenoids, resins, tannins and alkaloids. Bioactive compounds are secondary metabolites from trees and shrubs and are used in medicine, herbal medicine and cosmetology. To date, fruits and flowers of exotic trees and shrubs have been primarily used as sources of bioactive compounds. In turn, the search for new sources of bioactive compounds is currently focused on native plant species due to their availability. The application of such raw materials needs to be based on knowledge of their chemical composition, particularly health-promoting or therapeutic compounds. Research conducted to date on European trees and shrubs has been scarce. This paper presents the results of literature studies conducted to systematise the knowledge on phenolic compounds found in trees and shrubs native to central Europe. -

Hidden Fungi: Combining Culture-Dependent and -Independent DNA Barcoding Reveals Inter-Plant Variation in Species Richness of Endophytic Root Fungi in Elymus Repens
Journal of Fungi Article Hidden Fungi: Combining Culture-Dependent and -Independent DNA Barcoding Reveals Inter-Plant Variation in Species Richness of Endophytic Root Fungi in Elymus repens Anna K. Høyer and Trevor R. Hodkinson * Botany, School of Natural Sciences, Trinity College Dublin, The University of Dublin, Dublin D2, Ireland; [email protected] * Correspondence: [email protected] Abstract: The root endophyte community of the grass species Elymus repens was investigated using both a culture-dependent approach and a direct amplicon sequencing method across five sites and from individual plants. There was much heterogeneity across the five sites and among individual plants. Focusing on one site, 349 OTUs were identified by direct amplicon sequencing but only 66 OTUs were cultured. The two approaches shared ten OTUs and the majority of cultured endo- phytes do not overlap with the amplicon dataset. Media influenced the cultured species richness and without the inclusion of 2% MEA and full-strength MEA, approximately half of the unique OTUs would not have been isolated using only PDA. Combining both culture-dependent and -independent methods for the most accurate determination of root fungal species richness is therefore recom- mended. High inter-plant variation in fungal species richness was demonstrated, which highlights the need to rethink the scale at which we describe endophyte communities. Citation: Høyer, A.K.; Hodkinson, T.R. Hidden Fungi: Combining Culture-Dependent and -Independent Keywords: DNA barcoding; Elymus repens; fungal root endophytes; high-throughput amplicon DNA Barcoding Reveals Inter-Plant sequencing; MEA; PDA Variation in Species Richness of Endophytic Root Fungi in Elymus repens. J. Fungi 2021, 7, 466. -

Screening a Natural Product-Inspired Library for Anti-Phytophthora Activities
molecules Article Screening a Natural Product-Inspired Library for Anti-Phytophthora Activities Scott A. Lawrence 1, Hannah F. Robinson 2 , Daniel P. Furkert 3, Margaret A. Brimble 3 and Monica L. Gerth 2,* 1 Department of Microbiology and Immunology, University of Otago, Dunedin 9054, New Zealand; [email protected] 2 School of Biological Sciences, Victoria University of Wellington, Wellington 6140, New Zealand; [email protected] 3 School of Chemical Sciences, University of Auckland, Auckland 1142, New Zealand; [email protected] (D.P.F.); [email protected] (M.A.B.) * Correspondence: [email protected] Abstract: Phytophthora is a genus of microorganisms that cause devastating dieback and root-rot diseases in thousands of plant hosts worldwide. The economic impact of Phytophthora diseases on crops and native ecosystems is estimated to be billions of dollars per annum. These invasive pathogens are extremely difficult to control using existing chemical means, and the effectiveness of the few treatments available is being jeopardized by increasing rates of resistance. There is an urgent need to identify new chemical treatments that are effective against Phytophthora diseases. Natural products have long been regarded as “Nature’s medicine chest”, providing invaluable leads for developing front-line drugs and agrochemical agents. Here, we have screened a natural product- inspired library of 328 chemicals against two key Phytophthora species: Phytophthora cinnamomi and Phytophthora agathidicida. The library was initially screened for inhibition of zoospore germination. Citation: Lawrence, S.A.; Robinson, From these screens, we identified twenty-one hits that inhibited germination of one or both species.