Chapter 15 Plant Evolution and Classification Worksheets
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Gymnosperms the MESOZOIC: ERA of GYMNOSPERM DOMINANCE
Chapter 24 Gymnosperms THE MESOZOIC: ERA OF GYMNOSPERM DOMINANCE THE VASCULAR SYSTEM OF GYMNOSPERMS CYCADS GINKGO CONIFERS Pinaceae Include the Pines, Firs, and Spruces Cupressaceae Include the Junipers, Cypresses, and Redwoods Taxaceae Include the Yews, but Plum Yews Belong to Cephalotaxaceae Podocarpaceae and Araucariaceae Are Largely Southern Hemisphere Conifers THE LIFE CYCLE OF PINUS, A REPRESENTATIVE GYMNOSPERM Pollen and Ovules Are Produced in Different Kinds of Structures Pollination Replaces the Need for Free Water Fertilization Leads to Seed Formation GNETOPHYTES GYMNOSPERMS: SEEDS, POLLEN, AND WOOD THE ECOLOGICAL AND ECONOMIC IMPORTANCE OF GYMNOSPERMS The Origin of Seeds, Pollen, and Wood Seeds and Pollen Are Key Reproductive SUMMARY Innovations for Life on Land Seed Plants Have Distinctive Vegetative PLANTS, PEOPLE, AND THE Features ENVIRONMENT: The California Coast Relationships among Gymnosperms Redwood Forest 1 KEY CONCEPTS 1. The evolution of seeds, pollen, and wood freed plants from the need for water during reproduction, allowed for more effective dispersal of sperm, increased parental investment in the next generation and allowed for greater size and strength. 2. Seed plants originated in the Devonian period from a group called the progymnosperms, which possessed wood and heterospory, but reproduced by releasing spores. Currently, five lineages of seed plants survive--the flowering plants plus four groups of gymnosperms: cycads, Ginkgo, conifers, and gnetophytes. Conifers are the best known and most economically important group, including pines, firs, spruces, hemlocks, redwoods, cedars, cypress, yews, and several Southern Hemisphere genera. 3. The pine life cycle is heterosporous. Pollen strobili are small and seasonal. Each sporophyll has two microsporangia, in which microspores are formed and divide into immature male gametophytes while still retained in the microsporangia. -

"National List of Vascular Plant Species That Occur in Wetlands: 1996 National Summary."
Intro 1996 National List of Vascular Plant Species That Occur in Wetlands The Fish and Wildlife Service has prepared a National List of Vascular Plant Species That Occur in Wetlands: 1996 National Summary (1996 National List). The 1996 National List is a draft revision of the National List of Plant Species That Occur in Wetlands: 1988 National Summary (Reed 1988) (1988 National List). The 1996 National List is provided to encourage additional public review and comments on the draft regional wetland indicator assignments. The 1996 National List reflects a significant amount of new information that has become available since 1988 on the wetland affinity of vascular plants. This new information has resulted from the extensive use of the 1988 National List in the field by individuals involved in wetland and other resource inventories, wetland identification and delineation, and wetland research. Interim Regional Interagency Review Panel (Regional Panel) changes in indicator status as well as additions and deletions to the 1988 National List were documented in Regional supplements. The National List was originally developed as an appendix to the Classification of Wetlands and Deepwater Habitats of the United States (Cowardin et al.1979) to aid in the consistent application of this classification system for wetlands in the field.. The 1996 National List also was developed to aid in determining the presence of hydrophytic vegetation in the Clean Water Act Section 404 wetland regulatory program and in the implementation of the swampbuster provisions of the Food Security Act. While not required by law or regulation, the Fish and Wildlife Service is making the 1996 National List available for review and comment. -

The Glycosyltransferase Repertoire of the Spikemoss Selaginella Moellendorffiind a a Comparative Study of Its Cell Wall
Purdue University Purdue e-Pubs Department of Botany and Plant Pathology Faculty Department of Botany and Plant Pathology Publications 5-2-2012 The Glycosyltransferase Repertoire of the Spikemoss Selaginella moellendorffiind a a Comparative Study of Its Cell Wall. Jesper Harholt Iben Sørensen Cornell University Jonatan Fangel Alison Roberts University of Rhode Island William G.T. Willats See next page for additional authors Follow this and additional works at: http://docs.lib.purdue.edu/btnypubs Recommended Citation Harholt, Jesper; Sørensen, Iben; Fangel, Jonatan; Roberts, Alison; Willats, William G.T.; Scheller, Henrik Vibe; Petersen, Bent Larsen; Banks, Jo Ann; and Ulvskov, Peter, "The Glycosyltransferase Repertoire of the Spikemoss Selaginella moellendorffii and a Comparative Study of Its Cell Wall." (2012). Department of Botany and Plant Pathology Faculty Publications. Paper 15. http://dx.doi.org/10.1371/journal.pone.0035846 This document has been made available through Purdue e-Pubs, a service of the Purdue University Libraries. Please contact [email protected] for additional information. Authors Jesper Harholt, Iben Sørensen, Jonatan Fangel, Alison Roberts, William G.T. Willats, Henrik Vibe Scheller, Bent Larsen Petersen, Jo Ann Banks, and Peter Ulvskov This article is available at Purdue e-Pubs: http://docs.lib.purdue.edu/btnypubs/15 The Glycosyltransferase Repertoire of the Spikemoss Selaginella moellendorffii and a Comparative Study of Its Cell Wall Jesper Harholt1, Iben Sørensen1,2, Jonatan Fangel1, Alison Roberts3, William G. -

Prescribed Fire Decreases Lichen and Bryophyte Biomass and Alters Functional Group Composition in Pacific Northwest Prairies Author(S): Lalita M
Prescribed Fire Decreases Lichen and Bryophyte Biomass and Alters Functional Group Composition in Pacific Northwest Prairies Author(s): Lalita M. Calabria, Kate Petersen, Sarah T. Hamman and Robert J. Smith Source: Northwest Science, 90(4):470-483. Published By: Northwest Scientific Association DOI: http://dx.doi.org/10.3955/046.090.0407 URL: http://www.bioone.org/doi/full/10.3955/046.090.0407 BioOne (www.bioone.org) is a nonprofit, online aggregation of core research in the biological, ecological, and environmental sciences. BioOne provides a sustainable online platform for over 170 journals and books published by nonprofit societies, associations, museums, institutions, and presses. Your use of this PDF, the BioOne Web site, and all posted and associated content indicates your acceptance of BioOne’s Terms of Use, available at www.bioone.org/page/terms_of_use. Usage of BioOne content is strictly limited to personal, educational, and non-commercial use. Commercial inquiries or rights and permissions requests should be directed to the individual publisher as copyright holder. BioOne sees sustainable scholarly publishing as an inherently collaborative enterprise connecting authors, nonprofit publishers, academic institutions, research libraries, and research funders in the common goal of maximizing access to critical research. Lalita M. Calabria1, Kate Petersen,The Evergreen State College, 2700 Evergreen Parkway NW, Olympia, Washington 98505 Sarah T. Hamman, The Center for Natural Lands Management, 120 Union Ave SE #215, Olympia, Washington 98501 and Robert J. Smith, Department of Botany and Plant Pathology, 2082 Cordley Hall, Oregon State University, Corvallis, Oregon 97331 Prescribed Fire Decreases Lichen and Bryophyte Biomass and Alters Functional Group Composition in Pacific Northwest Prairies Abstract The reintroduction of fire to Pacific Northwest prairies has been useful for removing non-native shrubs and supporting habitat for fire-adapted plant and animal species. -
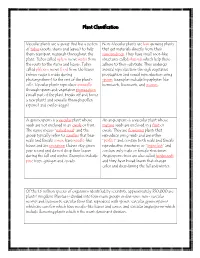
Plant Classification
Plant Classification Vascular plants are a group that has a system Non-Vascular plants are low growing plants of tubes (roots, stems and leaves) to help that get materials directly from their them transport materials throughout the surroundings. They have small root-like plant. Tubes called xylem move water from structures called rhizoids which help them the roots to the stems and leaves. Tubes adhere to their substrate. They undergo called phloem move food from the leaves asexual reproduction through vegetative (where sugar is made during propagation and sexual reproduction using photosynthesis) to the rest of the plant’s spores. Examples include bryophytes like cells. Vascular plants reproduce asexually hornworts, liverworts, and mosses. through spores and vegetative propagation (small part of the plant breaks off and forms a new plant) and sexually through pollen (sperm) and ovules (eggs). A gymnosperm is a vascular plant whose An angiosperm is a vascular plant whose seeds are not enclosed in an ovule or fruit. mature seeds are enclosed in a fruit or The name means “naked seed” and the ovule. They are flowering plants that group typically refers to conifers that bear reproduce using seeds and are either male and female cones, have needle-like “perfect” and contain both male and female leaves and are evergreen (leaves stay green reproductive structures or “imperfect” and year round and do not drop their leaves contain only male or female structures. during the fall and winter. Examples include Angiosperm trees are also called hardwoods pine trees, ginkgos and cycads. and they have broad leaves that change color and drop during the fall and winter. -

Bryophyte Life Cycle
MARCH 2016LEARNING | VELD & FLORA ABOUT BIODIVERSITY Veld & Flora MARCHFACTSHEET: 2016 | VELD MOSS & FLORA 24 25 3. BRYOPHYTE LIFE CYCLE Gametophyte (n) UNDERSTANDING THE GERMINATION MALE ALTERNATION OF GENERATIONS The way that almost all land plants reproduce is by means of two distinct, alternating life forms, a sexual phase that produces and releases gametes or sex cells and allows fertilisation, and a dispersal phase – both of which are adaptations to an essentially waterless environment. The sexual phase is known as the GAMETOPHYTE or haploid spores (n) generation and the dispersal phase is the SPOROPHYTE or diploid (2n) generation. Mature gametophyte plants produce haploid sex cells (egg and sperm) in sex organs (the male antheridia and female archegonia). These sex cells (also called gametes) fuse during fertilisation to form a diploid (2n) zygote which COPING OUT OF WATER grows, by means of mitosis (that results in two daughter cells each having Sperm (n) the same number and kind of chromosomes as the parent cell), into a new released Bryophytes, which include moss, are primitive plants that give us some idea sporophyte plant. The diploid sporophyte produces haploid (n) spores (i.e. from male of how the first plants that ventured onto land coped with their new waterless FEMALE each spore has a single set of chromosomes) by means of the process environment. They share many features with other plants, but differ in some of cell division called meiosis. Meiosis results in four daughter cells each ways – such as the lack of an effective vascular system (specialised tissue for with half the number of chromosomes of the parent cell. -

The Origin of Alternation of Generations in Land Plants
Theoriginof alternation of generations inlandplants: afocuson matrotrophy andhexose transport Linda K.E.Graham and LeeW .Wilcox Department of Botany,University of Wisconsin, 430Lincoln Drive, Madison,WI 53706, USA (lkgraham@facsta¡.wisc .edu ) Alifehistory involving alternation of two developmentally associated, multicellular generations (sporophyteand gametophyte) is anautapomorphy of embryophytes (bryophytes + vascularplants) . Microfossil dataindicate that Mid ^Late Ordovicianland plants possessed such alifecycle, and that the originof alternationof generationspreceded this date.Molecular phylogenetic data unambiguously relate charophyceangreen algae to the ancestryof monophyletic embryophytes, and identify bryophytes as early-divergentland plants. Comparison of reproduction in charophyceans and bryophytes suggests that the followingstages occurredduring evolutionary origin of embryophytic alternation of generations: (i) originof oogamy;(ii) retention ofeggsand zygotes on the parentalthallus; (iii) originof matrotrophy (regulatedtransfer ofnutritional and morphogenetic solutes fromparental cells tothe nextgeneration); (iv)origin of a multicellularsporophyte generation ;and(v) origin of non-£ agellate, walled spores. Oogamy,egg/zygoteretention andmatrotrophy characterize at least some moderncharophyceans, and arepostulated to represent pre-adaptativefeatures inherited byembryophytes from ancestral charophyceans.Matrotrophy is hypothesizedto have preceded originof the multicellularsporophytes of plants,and to represent acritical innovation.Molecular -

Spermatophyte Flora of Liangzi Lake Wetland Nature Reserve
E3S Web of Conferences 143, 02040 (2 020) https://doi.org/10.1051/e3sconf/20 2014302040 ARFEE 2019 Spermatophyte Flora of Liangzi Lake Wetland Nature Reserve Xinyang Zhang, Shijing He*, Rong Tao, and Huan Dai Wuhan Institute of Design and Sciences, Wuhan 430205, China Abstract. Based on route and sample-plot survey, plant resources of Liangzi Lake Wetland Nature Reserve were investigated. The result showed that there were 503 species of spermatophyte belonging to 296 genera of 86 families. There were 5 species under national first and second level protection. The dominant families of spermatophyte contained 20 species and above. The dominant genera of spermatophyte contained 4 species and below. The 86 families of spermatophyte can be divided into 7 distribution types and 4 variants. Tropic distribution type was dominant, accounting for 70.83% in total (excluding cosmopolitans). The 296 genera of spermatophyte can be divided into 14 distribution types and 9 variants. Temperate elements were a little more than tropical elements, accounting for 50.84% and 49.16% in total (excluding cosmopolitans) respectively. Reserve had 3 Chinese endemic genera, reflecting certain ancient and relict. The purpose of the research is to provide background information and scientific basis for the protection, construction, management and rational utilization of plant resources in the reserve. 1 Preface temperature is 17℃, the annual average rainfall is 1663mm, the average sunshine hour is 2061 hours, and Flora refers to the sum of all plant species in a certain the frost free period is 270 days. The rain bearing area is region or country. It is the result of the development and 208500 hectares, and the annual average water level is evolution of the plant kingdom under certain natural and 17.81m. -

Extant Diversity of Bryophytes Emerged from Successive Post-Mesozoic Diversification Bursts
ARTICLE Received 20 Mar 2014 | Accepted 3 Sep 2014 | Published 27 Oct 2014 DOI: 10.1038/ncomms6134 Extant diversity of bryophytes emerged from successive post-Mesozoic diversification bursts B. Laenen1,2, B. Shaw3, H. Schneider4, B. Goffinet5, E. Paradis6,A.De´samore´1,2, J. Heinrichs7, J.C. Villarreal7, S.R. Gradstein8, S.F. McDaniel9, D.G. Long10, L.L. Forrest10, M.L. Hollingsworth10, B. Crandall-Stotler11, E.C. Davis9, J. Engel12, M. Von Konrat12, E.D. Cooper13, J. Patin˜o1, C.J. Cox14, A. Vanderpoorten1,* & A.J. Shaw3,* Unraveling the macroevolutionary history of bryophytes, which arose soon after the origin of land plants but exhibit substantially lower species richness than the more recently derived angiosperms, has been challenged by the scarce fossil record. Here we demonstrate that overall estimates of net species diversification are approximately half those reported in ferns and B30% those described for angiosperms. Nevertheless, statistical rate analyses on time- calibrated large-scale phylogenies reveal that mosses and liverworts underwent bursts of diversification since the mid-Mesozoic. The diversification rates further increase in specific lineages towards the Cenozoic to reach, in the most recently derived lineages, values that are comparable to those reported in angiosperms. This suggests that low diversification rates do not fully account for current patterns of bryophyte species richness, and we hypothesize that, as in gymnosperms, the low extant bryophyte species richness also results from massive extinctions. 1 Department of Conservation Biology and Evolution, Institute of Botany, University of Lie`ge, Lie`ge 4000, Belgium. 2 Institut fu¨r Systematische Botanik, University of Zu¨rich, Zu¨rich 8008, Switzerland. -

Bryophyte Diversity and Vascular Plants
DISSERTATIONES BIOLOGICAE UNIVERSITATIS TARTUENSIS 75 BRYOPHYTE DIVERSITY AND VASCULAR PLANTS NELE INGERPUU TARTU 2002 DISSERTATIONES BIOLOGICAE UNIVERSITATIS TARTUENSIS 75 DISSERTATIONES BIOLOGICAE UNIVERSITATIS TARTUENSIS 75 BRYOPHYTE DIVERSITY AND VASCULAR PLANTS NELE INGERPUU TARTU UNIVERSITY PRESS Chair of Plant Ecology, Department of Botany and Ecology, University of Tartu, Estonia The dissertation is accepted for the commencement of the degree of Doctor philosophiae in plant ecology at the University of Tartu on June 3, 2002 by the Council of the Faculty of Biology and Geography of the University of Tartu Opponent: Ph.D. H. J. During, Department of Plant Ecology, the University of Utrecht, Utrecht, The Netherlands Commencement: Room No 218, Lai 40, Tartu on August 26, 2002 © Nele Ingerpuu, 2002 Tartu Ülikooli Kirjastuse trükikoda Tiigi 78, Tartu 50410 Tellimus nr. 495 CONTENTS LIST OF PAPERS 6 INTRODUCTION 7 MATERIAL AND METHODS 9 Study areas and field data 9 Analyses 10 RESULTS 13 Correlation between bryophyte and vascular plant species richness and cover in different plant communities (I, II, V) 13 Environmental factors influencing the moss and field layer (II, III) 15 Effect of vascular plant cover on the growth of bryophytes in a pot experiment (IV) 17 The distribution of grassland bryophytes and vascular plants into different rarity forms (V) 19 Results connected with nature conservation (I, II, V) 20 DISCUSSION 21 CONCLUSIONS 24 SUMMARY IN ESTONIAN. Sammaltaimede mitmekesisus ja seosed soontaimedega. Kokkuvõte 25 < TÄNUSÕNAD. Acknowledgements 28 REFERENCES 29 PAPERS 33 2 5 LIST OF PAPERS The present thesis is based on the following papers which are referred to in the text by the Roman numerals. -

Curriculum Vitae
CURRICULUM VITAE ORCID ID: 0000-0003-0186-6546 Gar W. Rothwell Edwin and Ruth Kennedy Distinguished Professor Emeritus Department of Environmental and Plant Biology Porter Hall 401E T: 740 593 1129 Ohio University F: 740 593 1130 Athens, OH 45701 E: [email protected] also Courtesy Professor Department of Botany and PlantPathology Oregon State University T: 541 737- 5252 Corvallis, OR 97331 E: [email protected] Education Ph.D.,1973 University of Alberta (Botany) M.S., 1969 University of Illinois, Chicago (Biology) B.A., 1966 Central Washington University (Biology) Academic Awards and Honors 2018 International Organisation of Palaeobotany lifetime Honorary Membership 2014 Fellow of the Paleontological Society 2009 Distinguished Fellow of the Botanical Society of America 2004 Ohio University Distinguished Professor 2002 Michael A. Cichan Award, Botanical Society of America 1999-2004 Ohio University Presidential Research Scholar in Biomedical and Life Sciences 1993 Edgar T. Wherry Award, Botanical Society of America 1991-1992 Outstanding Graduate Faculty Award, Ohio University 1982-1983 Chairman, Paleobotanical Section, Botanical Society of America 1972-1973 University of Alberta Dissertation Fellow 1971 Paleobotanical (Isabel Cookson) Award, Botanical Society of America Positions Held 2011-present Courtesy Professor of Botany and Plant Pathology, Oregon State University 2008-2009 Visiting Senior Researcher, University of Alberta 2004-present Edwin and Ruth Kennedy Distinguished Professor of Environmental and Plant Biology, Ohio -

Havens for Wildlife
)"7&/4'038*-%-*'& 4FDUJPO# .PTTFT -JWFSXPSUTBOE'FSOT This sheet explains what mosses, liverworts and Liverworts ferns are, where they occur and guidelines on how Liverworts are similar to to care for them in a burial ground. mosses but tend to be The earliest land plants were related to ferns and leafy and are less common. mosses. They started life growing on the edge of lakes Liverworts are mostly found and rivers 400 million years ago. Today most ferns, in woodland and by streams mosses and liverworts still need to grow in moist or rivers but can also be found places. on shady stones and damp soil in burial grounds. They are With our damp climate many burial sites prove ideal for strange, distinctive looking ferns, mosses and liverworts, particularly in the western plants and worth looking at areas of Britain and Ireland. closely or photographing their Adder’s-tongue Fern intricate shapes and subtle colours. MOSSES AND LIVERWORTS Mosses and liverworts are known as FERNS ‘bryophytes’. They are small, green plants *OUIFDPPM XFUDMJNBUFPGUIF6,UIFSFBSFUZQFTPG which do not have !owers or seeds but fern. Look out for ferns on west or north facing walls produce spores instead. There are over in particular. The shady areas of burial grounds, and 1000 di"erent species of bryophyte in under trees, where grass cutters don’t reach, will also UIF6,BOEUIFZUFOEUPCFGPVOEJO be places where ferns can !ourish undisturbed. sheltered, damp places as most of them cannot survive drying out. Few mosses An amazing fact about ferns is that once established PSMJWFSXPSUTIBWF&OHMJTIOBNFTBOEB they can survive in quite dry places, such as walls.