Taxonomy and Pylogeny of Symphytum with Special Emphasis
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

The Maiwa Guide to NATURAL DYES W H at T H Ey a R E a N D H Ow to U S E T H E M
the maiwa guide to NATURAL DYES WHAT THEY ARE AND HOW TO USE THEM WA L NUT NATURA L I ND IG O MADDER TARA SYM PL O C OS SUMA C SE Q UO I A MAR IG O L D SA FFL OWER B U CK THORN LIVI N G B L UE MYRO B A L AN K AMA L A L A C I ND IG O HENNA H I MA L AYAN RHU B AR B G A LL NUT WE L D P OME G RANATE L O G WOOD EASTERN B RA ZIL WOOD C UT C H C HAMOM IL E ( SA PP ANWOOD ) A LK ANET ON I ON S KI NS OSA G E C HESTNUT C O C H I NEA L Q UE B RA C HO EU P ATOR I UM $1.00 603216 NATURAL DYES WHAT THEY ARE AND HOW TO USE THEM Artisans have added colour to cloth for thousands of years. It is only recently (the first artificial dye was invented in 1857) that the textile industry has turned to synthetic dyes. Today, many craftspeople are rediscovering the joy of achieving colour through the use of renewable, non-toxic, natural sources. Natural dyes are inviting and satisfying to use. Most are familiar substances that will spark creative ideas and widen your view of the world. Try experimenting. Colour can be coaxed from many different sources. Once the cloth or fibre is prepared for dyeing it will soak up the colour, yielding a range of results from deep jew- el-like tones to dusky heathers and pastels. -

Tree of Life Marula Oil in Africa
HerbalGram 79 • August – October 2008 HerbalGram 79 • August Herbs and Thyroid Disease • Rosehips for Osteoarthritis • Pelargonium for Bronchitis • Herbs of the Painted Desert The Journal of the American Botanical Council Number 79 | August – October 2008 Herbs and Thyroid Disease • Rosehips for Osteoarthritis • Pelargonium for Bronchitis • Herbs of the Painted Desert • Herbs of the Painted Bronchitis for Osteoarthritis Disease • Rosehips for • Pelargonium Thyroid Herbs and www.herbalgram.org www.herbalgram.org US/CAN $6.95 Tree of Life Marula Oil in Africa www.herbalgram.org Herb Pharm’s Botanical Education Garden PRESERVING THE FULL-SPECTRUM OF NATURE'S CHEMISTRY The Art & Science of Herbal Extraction At Herb Pharm we continue to revere and follow the centuries-old, time- proven wisdom of traditional herbal medicine, but we integrate that wisdom with the herbal sciences and technology of the 21st Century. We produce our herbal extracts in our new, FDA-audited, GMP- compliant herb processing facility which is located just two miles from our certified-organic herb farm. This assures prompt delivery of freshly-harvested herbs directly from the fields, or recently HPLC chromatograph showing dried herbs directly from the farm’s drying loft. Here we also biochemical consistency of 6 receive other organic and wildcrafted herbs from various parts of batches of St. John’s Wort extracts the USA and world. In producing our herbal extracts we use precision scientific instru- ments to analyze each herb’s many chemical compounds. However, You’ll find Herb Pharm we do not focus entirely on the herb’s so-called “active compound(s)” at fine natural products and, instead, treat each herb and its chemical compounds as an integrated whole. -

Anchusa L. and Allied Genera (Boraginaceae) in Italy
Plant Biosystems - An International Journal Dealing with all Aspects of Plant Biology Official Journal of the Societa Botanica Italiana ISSN: 1126-3504 (Print) 1724-5575 (Online) Journal homepage: http://www.tandfonline.com/loi/tplb20 Anchusa L. and allied genera (Boraginaceae) in Italy F. SELVI & M. BIGAZZI To cite this article: F. SELVI & M. BIGAZZI (1998) Anchusa L. and allied genera (Boraginaceae) in Italy, Plant Biosystems - An International Journal Dealing with all Aspects of Plant Biology, 132:2, 113-142, DOI: 10.1080/11263504.1998.10654198 To link to this article: http://dx.doi.org/10.1080/11263504.1998.10654198 Published online: 18 Mar 2013. Submit your article to this journal Article views: 29 View related articles Citing articles: 20 View citing articles Full Terms & Conditions of access and use can be found at http://www.tandfonline.com/action/journalInformation?journalCode=tplb20 Download by: [Università di Pisa] Date: 05 November 2015, At: 02:31 PLANT BIOSYSTEMS, 132 (2) 113-142, 1998 Anchusa L. and allied genera (Boraginaceae) in Italy F. SEL VI and M. BIGAZZI received 18 May 1998; revised version accepted 30 July 1998 ABSTRACT - A revision of the Italian entities of Anchusa and of the rdated genera Anchusella, Lycopsis, Cynoglottis, Hormuzakia and Pentaglottis was carried out in view of the poor systematic knowledge of some entities of the national flora. The taxonomic treatment relies on a wide comparative basis, including macro- and micromorphological, karyological, chorological and ecological data. After a general description of some poorly known microCharacters of vegetative and reproductive structures, analytical keys, nomenclatural types, synonymies, descriptions, distribution maps and iconographies are provided for each entity. -

Gyneceum Apokarpní, Plodolistů Mnoho Na Prodlouženém Květním Lůžku
Magnoliaceae – šácholánovité Oddělení: Angiospermae (Magnoliophyta) Bazální krytosemenné Magnoliidy Řád: Magnoliales Čeleď: Magnoliaceae Magnoliaceae – šácholánovité • Stromy nebo keře (vždyzelené nebo opadavé) Liriodendron tulipifera Magnolia x soulangeana wihort.uwex.edu floralimages.co.uk Magnoliaceae – šácholánovité •Listy jednoduché, střídavé, s opadavými palisty Magnolia x soulangeana Liriodendron tulipifera everettj.people.cofc.edu wihort.uwex.edu Magnoliaceae – šácholánovité •Květy jednotlivé, většinou oboupohlavné, spirální •Květní obaly nerozlišené, tvoří okvětí Magnolia stellata Magnolia x soulangeana wihort.uwex.edu hcp4.net everettj.people.cofc.edu Liriodendron tulipifera floralimages.co.uk semillista.com Magnoliaceae – šácholánovité •Tyčinek mnoho, většinou lupenitých (krátká a tlustá nitka) •Gyneceum apokarpní, plodolistů mnoho na prodlouženém květním lůžku Liriodendron tulipifera nitky tyčinky pestíky semillista.com semillista.com Magnoliaceae – šácholánovité •Plod měchýřek (Magnolia), nažka (Liriodendron), tvoří šišticovitá souplodí •častá kantarogamie (opylení brouky) Magnolia grandiflora Liriodendron tulipifera commons.wikimedia.org Primulaceae (prvosenkovité) Oddělení: Angiospermae (Magnoliophyta) Dvouděložné (Dicots) Pravé dvouděložné (Eudicots) Asteridy Řád: Ericales Čeleď: Primulaceae Primulaceae – prvosenkovité • Převážně suchozemské byliny, vzácně vodní rostliny okolík květů • Listy střídavé, vstřícné nebo v přízemní růžici většinou jednoduché, u vodních složené bez palistů • Květy jednotlivé nebo v květenstvích -

Anchusa Crispa Viv. Subsp. Crispa
IN SITU CONSERVATION - SPECIES INFORMATION SHEET Project partner: National Botanical Conservatory of Corsica of the Office of the Environment of Corsica Island CORSICA Species name (Family) Anchusa crispa Viv. subsp. crispa (Boraginaceae) Common name French: Buglosse crépue Plant description V Herbaceous taxa, hispid, initially in a rosette of hard leaves, prostrate on the sand, then presenting with floriferous stems, leafy, littlely branched from the rosette. Leaved leaves, oblong, sinuated-crispy, from 4-10 mm broad to 3-20 cm long. Terminal inflorescences fomed uniparous scorpioid cymes, elongating during the flowering period, with bearing flowers onaxils of small hairy bracts. Small, actinomorphous, gamosepal calyx flowers and blue gamopetal corolla. Fruits: 4 achenes per flower, with a very small aril. V Flowering occurs between April and May. Fruiting from late May to September. Pollination is done by insects and the seed spread is generally barochorial, even some cases of zoochory is signaled due to ants and caws. V A. crispa is a sabulicolous, heliophilous, nitrophilous or subnitrophilic plant, xerophilic, well tolerant of sea salt, but very sensitive to sand cover during storms Distribution This species is endemic of Corsica and Sardinia. There are: 2 populations located in the east coast of Corsica and 8 populations located on the sand along the North West of Sardinia. Map Two subspecies of A. crispa exist in Corsica: A. crispa valincoana and A. crispa crispa . The crispa subspecies is only present on the east coast of the island. There are only two populations highly threatened by tourism activities. IN SITU CONSERVATION - SPECIES INFORMATION SHEET Legal status A. -

State of New York City's Plants 2018
STATE OF NEW YORK CITY’S PLANTS 2018 Daniel Atha & Brian Boom © 2018 The New York Botanical Garden All rights reserved ISBN 978-0-89327-955-4 Center for Conservation Strategy The New York Botanical Garden 2900 Southern Boulevard Bronx, NY 10458 All photos NYBG staff Citation: Atha, D. and B. Boom. 2018. State of New York City’s Plants 2018. Center for Conservation Strategy. The New York Botanical Garden, Bronx, NY. 132 pp. STATE OF NEW YORK CITY’S PLANTS 2018 4 EXECUTIVE SUMMARY 6 INTRODUCTION 10 DOCUMENTING THE CITY’S PLANTS 10 The Flora of New York City 11 Rare Species 14 Focus on Specific Area 16 Botanical Spectacle: Summer Snow 18 CITIZEN SCIENCE 20 THREATS TO THE CITY’S PLANTS 24 NEW YORK STATE PROHIBITED AND REGULATED INVASIVE SPECIES FOUND IN NEW YORK CITY 26 LOOKING AHEAD 27 CONTRIBUTORS AND ACKNOWLEGMENTS 30 LITERATURE CITED 31 APPENDIX Checklist of the Spontaneous Vascular Plants of New York City 32 Ferns and Fern Allies 35 Gymnosperms 36 Nymphaeales and Magnoliids 37 Monocots 67 Dicots 3 EXECUTIVE SUMMARY This report, State of New York City’s Plants 2018, is the first rankings of rare, threatened, endangered, and extinct species of what is envisioned by the Center for Conservation Strategy known from New York City, and based on this compilation of The New York Botanical Garden as annual updates thirteen percent of the City’s flora is imperiled or extinct in New summarizing the status of the spontaneous plant species of the York City. five boroughs of New York City. This year’s report deals with the City’s vascular plants (ferns and fern allies, gymnosperms, We have begun the process of assessing conservation status and flowering plants), but in the future it is planned to phase in at the local level for all species. -

FDA OTC Reviews Summary of Back Issues
Number 23 The Journal of the AMERICAN BOTANI CAL COUNCIL and the HERB RESEARCH FOUNDATION Chinese Medicinals -A Comprehensive Review of Chinese Materia Medica Legal and Regulatory- FDA OTC Reviews Summary of Back Issues Ongoing Market Report, Research Reviews (glimpses of studies published in over a dozen scientific and technical journals), Access, Book Reviews, Calendar, Legal and Regulatory, Herb Blurbs and Potpourri columns. #1 -Summer 83 (4 pp.) Eucalyptus Repels Reas, Stones Koalas; FDA OTC tiveness; Fungal Studies; More Polysaccharides; Recent Research on Ginseng; Heart Panel Reviews Menstrual & Aphrodisiac Herbs; Tabasco Toxicity?; Garlic Odor Peppers; Yew Continues to Amaze; Licorice O.D. Prevention; Ginseng in Perspec Repels Deer; and more. tive; Poisonous Plants Update; Medicinal Plant Conservation Project; 1989 Oberly #2- Fall/Winter 83-84 (8 pp.) Appeals Court Overrules FDA on Food Safety; Award Nominations; Trends in Self-Care Conference; License Plates to Fund Native FDA Magazine Pans Herbs; Beware of Bay Leaves; Tiny Tree: Cancer Cure?; Plant Manual; and more. Comfrey Tea Recall; plus. #17-Summer 88. (24 pp.) Sarsaparilla, A Literature Review by Christopher #3-Spring 84 (8 pp.) Celestial Sells to Kraft; Rowers and Dinosaurs Demise?; Hobbs; Hops May Help Metabolize Toxins; Herbal Roach Killer; Epazote Getting Citrus Peels for Kitty Litter; Saffron; Antibacterial Sassafras; WHO Studies Anti· More Popular, Aloe Market Levels Off; Herbal Tick Repellent?; Chinese Herb fertility Plants; Chinese Herbal Drugs; Feverfew Migraines; -

Diplomová Práce
Jihočeská univerzita v Českých Budějovicích Zemědělská fakulta Studijní program: N4101 Zemědělské inženýrství Studijní obor: Agroekologie Katedra: Rostlinné výroby a agroekologie Vedoucí katedry: prof. Ing. Vladislav Čurn, Ph.D. DIPLOMOVÁ PRÁCE Biologie, semenářské vlastnosti a zakládání porostů kostivalu lékařského (Symphytum officinale L.) Vedoucí diplomové práce: Ing. Milan Kobes, Ph.D. Autor diplomové práce: Bc. Jaroslava Kovářová České Budějovice, duben 2016 Čestné prohlášení Prohlašuji, že svou diplomovou práci s názvem „Biologie, semenářské vlastnosti a zakládání porostů kostivalu lékařského (Symphytum officinale L.)“ jsem vypracovala samostatně pod vedením vedoucí diplomové práce Ing. Milana Kobese, Ph.D a s použitím odborné literatury a dalších informačních zdrojů, které jsou v práci citovány a zároveň uvedeny v seznamu literatury na konci této práce. Jako autorka uvedené diplomové práce dále prohlašuji, že jsem v souvislosti s jejím vytvořením neporušila autorská práva třetích osob. Dále prohlašuji, že v souladu s § 47b zákona č. 111/1998 Sb. v platném znění souhlasím se zveřejněním své diplomové práce, a to v nezkrácené podobě (v úpravě vzniklé vypuštěním vyznačených částí archivovaných Zemědělskou fakultou JU) elektronickou cestou ve veřejně přístupné části databáze STAG provozované Jihočeskou univerzitou v Českých Budějovicích na jejích internetových stránkách. V Českých Budějovicích dne.......... _____________________________ Poděkování Velké poděkování patří mému vedoucímu práce Ing. Milanu Kobesovi, Ph.D. za odborné rady, připomínky, za vedení a odborné konzultace, které mi poskytl během zpracování této diplomové práce. Děkuji celé své rodině za podporu. ABSTRAKT Diplomová práce s názvem ,,Biologie, semenářské vlastnosti a zakládání porostů kostivalu lékařského (Symphytum officinale L.)“ se zabývala hodnocením způsobů rozmnožování kostivalu a hodnocením vlivu ekologických podmínek na růst kostivalu. Dále testy klíčivosti a jejich statistickým hodnocením při respektování specifik práce se semeny. -
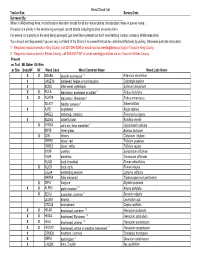
Weed Check List Survey Date
Weed Check List Trail or Site: Survey Date: Surveyed By: When in Wildnerness Area, record location and other details for all non-native plants. Include plant name in survey notes. If a weed is a priority in the area being surveyed, record details including location on survey form. For weeds not a priority in the area being surveyed, just mark them present but don't need lat/long location unless in Wilderness Area. You can pull and bag weeds if you are very confident of the ID and it is a weed that can be controlled effectively by pulling. Otherwise just note its location. 1 - Regulated noxious weeds in King County, call 206-296-0290 or email [email protected] if found in King County 2 - Regulated noxious weed in Kittitas County, call 509-962-7007 or email [email protected] if found in Kittitas County Present on Trail Mt. Baker Ok Wen or Site Snoq NF NF Weed Code Weed Common Name Weed Latin Name XXARAB3 absinth wormwood 1,2 Artemisia absinthium X CASE13 bindweed, hedge or morning glory Calystegia sepium X SODU bittersweet nightshade Solanum dulcamara XXRULA blackberry, evergreen or cutleaf 2 Rubus laciniatus XXRUAR9 blackberry, Himalayan 2 Rubus armeniacus SILA21 bladder campion 2 Silene latifolia X AJRE bugleweed Ajuga reptans RARE3 buttercup, creeping Ranunculus repens X BUDA2 butterfly bush Buddleia davidii X HYRA3 cat’s ear, false dandelion 2 Hypochaeris radicata BRTE cheat grass Bromus tectorum X CIIN chicory Cichorium intybus TRPR2 clover, red Trifolium pratense TRRE3 clover, white Trifolium repens SYOF comfrey Symphytum officinale TAOF dandelion Taraxacum officinale RUOB dock, broadleaf Rumex obtusifolius X RUCR dock, curly Rumex crispus LALA4 everlasting peavine Lathyrus latifolius MAPE2 false mayweed Tripleurospermum perforatum X DIPU foxglove Digitalis purpurea XXALPE4 garlic mustard 1,2 Alliaria petiolata X SEVU groundsel, common 2 Senecio vulgaris LEONT hawkbit Leontodon spp. -

Two New Genera in the Omphalodes Group (Cynoglosseae, Boraginaceae)
Nova Acta Científica Compostelana (Bioloxía),23 : 1-14 (2016) - ISSN 1130-9717 ARTÍCULO DE INVESTIGACIÓN Two new genera in the Omphalodes group (Cynoglosseae, Boraginaceae) Dous novos xéneros no grupo Omphalodes (Cynoglosseae, Boraginaceae) M. SERRANO1, R. CARBAJAL1, A. PEREIRA COUTINHO2, S. ORTIZ1 1 Department of Botany, Faculty of Pharmacy, University of Santiago de Compostela, 15782 Santiago de Compostela , Spain 2 CFE, Centre for Functional Ecology, Department of Life Sciences, University of Coimbra, 3000-456 Coimbra, Portugal *[email protected]; [email protected]; [email protected]; [email protected] *: Corresponding author (Recibido: 08/06/2015; Aceptado: 01/02/2016; Publicado on-line: 04/02/2016) Abstract Omphalodes (Boraginaceae, Cynoglosseae) molecular phylogenetic relationships are surveyed in the context of the tribe Cynoglosseae, being confirmed that genusOmphalodes is paraphyletic. Our work is focused both in the internal relationships among representatives of Omphalodes main subgroups (and including Omphalodes verna, the type species), and their relationships with other Cynoglosseae genera that have been related to the Omphalodes group. Our phylogenetic analysis of ITS and trnL-trnF molecular markers establish close relationships of the American Omphalodes with the genus Mimophytum, and also with Cynoglossum paniculatum and Myosotidium hortensia. The southwestern European annual Omphalodes species form a discrete group deserving taxonomic recognition. We describe two new genera to reduce the paraphyly in the genus Omphalodes, accommodating the European annual species in Iberodes and Cynoglossum paniculatum in Mapuchea. The pollen of the former taxon is described in detail for the first time. Keywords: Madrean-Tethyan, phylogeny, pollen, systematics, taxonomy Resumo Neste estudo analisamos as relacións filoxenéticas deOmphalodes (Boraginaceae, Cynoglosseae) no contexto da tribo Cynoglosseae, confirmándose como parafilético o xéneroOmphalodes . -

Brozura Alkana Anglicka Zlom.Indd
Text: Ing. Marta Mútňanová Ing. Libor Ulrych, PhD. Translati on: RNDr. Radoslav Považan, Mgr. Mária Gréčová Graphic: Ing. Viktória Ihringová Photographs by (see pictures): Ing. Marta Mútňanová (No. 1 – the plant, 2, 3, 4, 9, 12, 13, 14, 15, 17) Ing. Libor Ulrych, PhD. (No. 1 – detail of infl orescence, 5, 6, 7, 8, 11) RNDr. Alžbeta Szabóová (No. 10, 16) Published by the State Nature Conservancy of the Slovak Republic 2010 introduction In 2009 the State Nature Conservancy of the Slovak Republic applied for a non-re- fundable subsidy from the Ministry of Environment of the Slovak Republic within the Operati onal Programme Environment, priority axis 5. Protecti on and Regenerati on of Natural Environment and Landscape, in order to ensure implementati on of program- mes to preserve alkanet. Operati onal Programme Environment aims at use of fi nancial resources from the Euro- pean Regional Development Fund and the Cohesion Fund in the environment protecti - on in the period 2007 – 2013. Its objecti ve is to improve environmental status as well as the rati onal use of natural resources. The project enti tled Implementati on of programmes to preserve criti cally endange- red plant species was approved in September 2009. It focused on preservati on of two criti cally endangered plant species: alkanet – Alkanna ti nctoria and autumn crocus – Colchicum arenarium. The State Nature Conservancy of the Slovak Republic elabora- ted conservati on programmes for both species. On February 25, 2009 the programmes were approved by the Ministry of Environment at an operati ve session of the Minister of Environment in Decree No. -

ISTA List of Stabilized Plant Names 7Th Edition
ISTA List of Stabilized Plant Names th 7 Edition ISTA Nomenclature Committee Chair: Dr. M. Schori Published by All rights reserved. No part of this publication may be The Internation Seed Testing Association (ISTA) reproduced, stored in any retrieval system or transmitted Zürichstr. 50, CH-8303 Bassersdorf, Switzerland in any form or by any means, electronic, mechanical, photocopying, recording or otherwise, without prior ©2020 International Seed Testing Association (ISTA) permission in writing from ISTA. ISBN 978-3-906549-77-4 ISTA List of Stabilized Plant Names 1st Edition 1966 ISTA Nomenclature Committee Chair: Prof P. A. Linehan 2nd Edition 1983 ISTA Nomenclature Committee Chair: Dr. H. Pirson 3rd Edition 1988 ISTA Nomenclature Committee Chair: Dr. W. A. Brandenburg 4th Edition 2001 ISTA Nomenclature Committee Chair: Dr. J. H. Wiersema 5th Edition 2007 ISTA Nomenclature Committee Chair: Dr. J. H. Wiersema 6th Edition 2013 ISTA Nomenclature Committee Chair: Dr. J. H. Wiersema 7th Edition 2019 ISTA Nomenclature Committee Chair: Dr. M. Schori 2 7th Edition ISTA List of Stabilized Plant Names Content Preface .......................................................................................................................................................... 4 Acknowledgements ....................................................................................................................................... 6 Symbols and Abbreviations ..........................................................................................................................