PHYLOGENETIC and BIOGEOGRAPHIC PATTERNS of BASILISCINE IGUANIANS (Reptilia: Squamata: "Iguanidae")
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Checklist of Helminths from Lizards and Amphisbaenians (Reptilia, Squamata) of South America Ticle R A
The Journal of Venomous Animals and Toxins including Tropical Diseases ISSN 1678-9199 | 2010 | volume 16 | issue 4 | pages 543-572 Checklist of helminths from lizards and amphisbaenians (Reptilia, Squamata) of South America TICLE R A Ávila RW (1), Silva RJ (1) EVIEW R (1) Department of Parasitology, Botucatu Biosciences Institute, São Paulo State University (UNESP – Univ Estadual Paulista), Botucatu, São Paulo State, Brazil. Abstract: A comprehensive and up to date summary of the literature on the helminth parasites of lizards and amphisbaenians from South America is herein presented. One-hundred eighteen lizard species from twelve countries were reported in the literature harboring a total of 155 helminth species, being none acanthocephalans, 15 cestodes, 20 trematodes and 111 nematodes. Of these, one record was from Chile and French Guiana, three from Colombia, three from Uruguay, eight from Bolivia, nine from Surinam, 13 from Paraguay, 12 from Venezuela, 27 from Ecuador, 17 from Argentina, 39 from Peru and 103 from Brazil. The present list provides host, geographical distribution (with the respective biome, when possible), site of infection and references from the parasites. A systematic parasite-host list is also provided. Key words: Cestoda, Nematoda, Trematoda, Squamata, neotropical. INTRODUCTION The present checklist summarizes the diversity of helminths from lizards and amphisbaenians Parasitological studies on helminths that of South America, providing a host-parasite list infect squamates (particularly lizards) in South with localities and biomes. America had recent increased in the past few years, with many new records of hosts and/or STUDIED REGIONS localities and description of several new species (1-3). -

Iii Pontificia Universidad Católica Del
III PONTIFICIA UNIVERSIDAD CATÓLICA DEL ECUADOR FACULTAD DE CIENCIAS EXACTAS Y NATURALES ESCUELA DE CIENCIAS BIOLÓGICAS Un método integrativo para evaluar el estado de conservación de las especies y su aplicación a los reptiles del Ecuador Tesis previa a la obtención del título de Magister en Biología de la Conservación CAROLINA DEL PILAR REYES PUIG Quito, 2015 IV CERTIFICACIÓN Certifico que la disertación de la Maestría en Biología de la Conservación de la candidata Carolina del Pilar Reyes Puig ha sido concluida de conformidad con las normas establecidas; por tanto, puede ser presentada para la calificación correspondiente. Dr. Omar Torres Carvajal Director de la Disertación Quito, Octubre del 2015 V AGRADECIMIENTOS A Omar Torres-Carvajal, curador de la División de Reptiles del Museo de Zoología de la Pontificia Universidad Católica del Ecuador (QCAZ), por su continua ayuda y contribución en todas las etapas de este estudio. A Andrés Merino-Viteri (QCAZ) por su valiosa ayuda en la generación de mapas de distribución potencial de reptiles del Ecuador. A Santiago Espinosa y Santiago Ron (QCAZ) por sus acertados comentarios y correcciones. A Ana Almendáriz por haber facilitado las localidades geográficas de presencia de ciertos reptiles del Ecuador de la base de datos de la Escuela Politécnica Nacional (EPN). A Mario Yánez-Muñoz de la División de Herpetología del Museo Ecuatoriano de Ciencias Naturales del Instituto Nacional de Biodiversidad (DHMECN-INB), por su ayuda y comentarios a la evaluación de ciertos reptiles del Ecuador. A Marcio Martins, Uri Roll, Fred Kraus, Shai Meiri, Peter Uetz y Omar Torres- Carvajal del Global Assessment of Reptile Distributions (GARD) por su colaboración y comentarios en las encuestas realizadas a expertos. -

Iguanidae: Hoplocercinae) from Southwestern Ecuador
Volume 48(20):227-235, 2008 A new species of ENYALIOIDES (Iguanidae: Hoplocercinae) from southwestern Ecuador Omar Torres-Carvajal1,2 Ana Almendáriz3 Jorge Valencia4 Mario Yánez-Muñoz5 Juan P. Reyes5 ABStract We describe a new species of Enyalioides from lowland cloud forests in southwestern Ecuador. This represents the third species in the genus known to occur west of the Andes in South America; the other two species are E. heterolepis and E. oshaughnessyi. Among other characters, the new species can be distinguished from other members in the genus by having small, keeled, paravertebrals; a series of skin folds on the lateral aspects of body and neck; size-homogeneous scales on body and limbs; distinct caudal segments; and an extensive dark patch on the gular region of adult males. Morphological similarity suggests that the new species, which we call E. touzeti, is closely related to E. oshaughnessyi. Keywords: Ecuador, Enyalioides, Hoplocercinae, Iguania, new species. INtroDUctioN Panama and southeastern Brazil, on both sides of the Andes, with most species occurring in Colombia, Ec- The neotropical iguanian lizard clade Hoplocer- uador, and Peru (Table 1). cidae (Frost & Etheridge, 1989; Frost et al., 2001), Despite the small size of this clade and its phylo- also known as Hoplocercinae (Macey et al., 1997; genetic importance as a possible basal lineage within Schulte et al., 2003), includes 11 species in three Iguania (Etheridge & de Queiroz, 1988; Schulte et al., genera (Enyalioides, Hoplocercus, and Morunasaurus). 1998, 2003), many questions remain to be answered. Hoplocercines are known from the lowlands between First, the phylogenetic relationships among its species 1. -

Literature Cited in Lizards Natural History Database
Literature Cited in Lizards Natural History database Abdala, C. S., A. S. Quinteros, and R. E. Espinoza. 2008. Two new species of Liolaemus (Iguania: Liolaemidae) from the puna of northwestern Argentina. Herpetologica 64:458-471. Abdala, C. S., D. Baldo, R. A. Juárez, and R. E. Espinoza. 2016. The first parthenogenetic pleurodont Iguanian: a new all-female Liolaemus (Squamata: Liolaemidae) from western Argentina. Copeia 104:487-497. Abdala, C. S., J. C. Acosta, M. R. Cabrera, H. J. Villaviciencio, and J. Marinero. 2009. A new Andean Liolaemus of the L. montanus series (Squamata: Iguania: Liolaemidae) from western Argentina. South American Journal of Herpetology 4:91-102. Abdala, C. S., J. L. Acosta, J. C. Acosta, B. B. Alvarez, F. Arias, L. J. Avila, . S. M. Zalba. 2012. Categorización del estado de conservación de las lagartijas y anfisbenas de la República Argentina. Cuadernos de Herpetologia 26 (Suppl. 1):215-248. Abell, A. J. 1999. Male-female spacing patterns in the lizard, Sceloporus virgatus. Amphibia-Reptilia 20:185-194. Abts, M. L. 1987. Environment and variation in life history traits of the Chuckwalla, Sauromalus obesus. Ecological Monographs 57:215-232. Achaval, F., and A. Olmos. 2003. Anfibios y reptiles del Uruguay. Montevideo, Uruguay: Facultad de Ciencias. Achaval, F., and A. Olmos. 2007. Anfibio y reptiles del Uruguay, 3rd edn. Montevideo, Uruguay: Serie Fauna 1. Ackermann, T. 2006. Schreibers Glatkopfleguan Leiocephalus schreibersii. Munich, Germany: Natur und Tier. Ackley, J. W., P. J. Muelleman, R. E. Carter, R. W. Henderson, and R. Powell. 2009. A rapid assessment of herpetofaunal diversity in variously altered habitats on Dominica. -

Redalyc.Comparative Studies of Supraocular Lepidosis in Squamata
Multequina ISSN: 0327-9375 [email protected] Instituto Argentino de Investigaciones de las Zonas Áridas Argentina Cei, José M. Comparative studies of supraocular lepidosis in squamata (reptilia) and its relationships with an evolutionary taxonomy Multequina, núm. 16, 2007, pp. 1-52 Instituto Argentino de Investigaciones de las Zonas Áridas Mendoza, Argentina Disponible en: http://www.redalyc.org/articulo.oa?id=42801601 Cómo citar el artículo Número completo Sistema de Información Científica Más información del artículo Red de Revistas Científicas de América Latina, el Caribe, España y Portugal Página de la revista en redalyc.org Proyecto académico sin fines de lucro, desarrollado bajo la iniciativa de acceso abierto ISSN 0327-9375 COMPARATIVE STUDIES OF SUPRAOCULAR LEPIDOSIS IN SQUAMATA (REPTILIA) AND ITS RELATIONSHIPS WITH AN EVOLUTIONARY TAXONOMY ESTUDIOS COMPARATIVOS DE LA LEPIDOSIS SUPRA-OCULAR EN SQUAMATA (REPTILIA) Y SU RELACIÓN CON LA TAXONOMÍA EVOLUCIONARIA JOSÉ M. CEI † las subfamilias Leiosaurinae y RESUMEN Enyaliinae. Siempre en Iguania Observaciones morfológicas Pleurodonta se evidencian ejemplos previas sobre un gran número de como los inconfundibles patrones de especies permiten establecer una escamas supraoculares de correspondencia entre la Opluridae, Leucocephalidae, peculiaridad de los patrones Polychrotidae, Tropiduridae. A nivel sistemáticos de las escamas específico la interdependencia en supraoculares de Squamata y la Iguanidae de los géneros Iguana, posición evolutiva de cada taxón Cercosaura, Brachylophus, -

A Checklist of the Amphibians and Reptiles of Honduras, with Additions, Comments on Taxonomy, Some Recent Taxonomic Decisions, and Areas of Further Studies Needed
Zootaxa 3931 (3): 352–386 ISSN 1175-5326 (print edition) www.mapress.com/zootaxa/ Article ZOOTAXA Copyright © 2015 Magnolia Press ISSN 1175-5334 (online edition) http://dx.doi.org/10.11646/zootaxa.3931.3.2 http://zoobank.org/urn:lsid:zoobank.org:pub:93296D90-0DF4-4FD3-A63F-8354EC89D40C A checklist of the amphibians and reptiles of Honduras, with additions, comments on taxonomy, some recent taxonomic decisions, and areas of further studies needed JAMES R. MCCRANIE 10770 SW 164 Street, Miami, Florida, 33157–2933, USA. E-mail: [email protected] Abstract An updated checklist of the amphibians and reptiles of Honduras is provided. The list includes three amphibian species (Ptychohyla euthysanota, Bolitoglossa odonnelli, Oedipina chortiorum) and two reptile species (Laemanctus waltersi [el- evated from subspecies status], Epictia phenops) not included in the most recent checklist of the amphibians and reptiles of the country. Also, one amphibian genus and species (Triprion petasatus) is removed from the country list and one Hon- duran lizard (Ctenosaura praeocularis) is synonymized with an older name. Comments where more study is needed are also included where pertinent. Authors, dates, and original spellings of the higher-level taxonomy of all taxa covered here- in are also given. A total of 401 species (137 amphibians and 264 reptiles) are now known from the country with 111 spe- cies (27.7%) being Honduran endemics (52 amphibians and 59 reptiles). Key words: Amphibians, reptiles, taxonomic checklist, Honduras, Central America Introduction Seven countries make up political Central America. Of those seven countries, Honduras is among the leaders in the amount of progress being made towards an understanding of its amphibian and reptile diversity. -

First Record of Oviposition Event by Laemanctus Longipes (Sauria: Corytophanidae) in Mexico
SHORT NOTE https://dx.doi.org/10.15446/caldasia.v40n2.68091 http://www.revistas.unal.edu.co/index.php/cal Caldasia 40(2):390-393. Julio-diciembre 2018 First record of oviposition event by Laemanctus longipes (Sauria: Corytophanidae) in Mexico Primer reporte de puesta de huevos de Laemanctus longipes (Sauria: Corytophanidae) en México ARTURO GONZÁLEZ-ZAMORA1*, SÒNIA SÁNCHEZ-LÓPEZ2, CHRISTIAN A. DELFÍN-ALFONSO1, E. AHMED BELLO-SÁNCHEZ3 1Instituto de Investigaciones Biológicas, Universidad Veracruzana, Xalapa, Veracruz, Mexico. [email protected]*, [email protected] 2Dirección del Área Biológico-Agropecuarias, Universidad Veracruzana, Xalapa, Veracruz, Mexico. [email protected] 3Instituto de Neuroetología, Universidad Veracruzana, Xalapa, Veracruz, Mexico. [email protected] * Corresponding author. ABSTRACT Oviposition in elusive reptiles has been poorly documented due to the difficulty in observing it in natural habitats. Here, we document the first egg-laying record in the Eastern Casque- headed Basilisk Laemanctus longipes in the wild. Our record adds novel information about the oviposition of this scarcely studied reptile species. Key words. Egg laying, elusive species, Eastern Casque-headed Basilisk, smooth casque- headed basilisk. RESUMEN La oviposición en especies elusivas de reptiles ha sido escasamente registrada en vida libre debido a la dificultad para observarla. En este trabajo documentamos por primera vez un evento de puesta de huevos en vida libre del toloque verde Laemanctus longipes en el centro del estado de Veracruz, México. Este registro agrega información novedosa sobre la oviposición de esta especie de reptil poco estudiada. Palabras clave. Puesta de huevos, especies elusivas, toloque verde, lemacto coludo. Oviposition behavior in elusive reptile and Central America, Laemanctus species species has been poorly recorded due to the present restricted geographical distributions difficulty of field observation (Campbell (Köhler 2008). -

A Phylogeny and Revised Classification of Squamata, Including 4161 Species of Lizards and Snakes
BMC Evolutionary Biology This Provisional PDF corresponds to the article as it appeared upon acceptance. Fully formatted PDF and full text (HTML) versions will be made available soon. A phylogeny and revised classification of Squamata, including 4161 species of lizards and snakes BMC Evolutionary Biology 2013, 13:93 doi:10.1186/1471-2148-13-93 Robert Alexander Pyron ([email protected]) Frank T Burbrink ([email protected]) John J Wiens ([email protected]) ISSN 1471-2148 Article type Research article Submission date 30 January 2013 Acceptance date 19 March 2013 Publication date 29 April 2013 Article URL http://www.biomedcentral.com/1471-2148/13/93 Like all articles in BMC journals, this peer-reviewed article can be downloaded, printed and distributed freely for any purposes (see copyright notice below). Articles in BMC journals are listed in PubMed and archived at PubMed Central. For information about publishing your research in BMC journals or any BioMed Central journal, go to http://www.biomedcentral.com/info/authors/ © 2013 Pyron et al. This is an open access article distributed under the terms of the Creative Commons Attribution License (http://creativecommons.org/licenses/by/2.0), which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited. A phylogeny and revised classification of Squamata, including 4161 species of lizards and snakes Robert Alexander Pyron 1* * Corresponding author Email: [email protected] Frank T Burbrink 2,3 Email: [email protected] John J Wiens 4 Email: [email protected] 1 Department of Biological Sciences, The George Washington University, 2023 G St. -

Amphibians and Reptiles
Chapter 11 Amphibians and Reptiles Pierre Charruau, Jose´ Rogelio Cedeno-Va~ ´zquez, and Gunther Kohler€ Abstract The three Mexican states of the Yucata´n Peninsula have been relatively well explored for herpetofauna, when compared with other states of the country. However, most studies on the herpetofauna of the Yucata´n Peninsula have focused on their diversity, taxonomy, and species distribution, and less on their ecology, behavior or conservation status. The major conservation efforts have focused on sea turtles. Although some conservation programs exist locally for crocodiles in the north of the peninsula, to date conservation strategies have mostly been restricted to the designation of protected areas. With 24 species of amphibians and 118 species of reptiles, the Yucata´n Peninsula harbors 11.5 % of national herpetofauna diver- sity, and 19 % of species are endemic to the peninsula. Reptiles and amphibians are two major globally threatened groups of vertebrates, with amphibians being the most threatened vertebrate class. Both groups face the same threats, namely habitat loss and modification, pollution, overharvest for food and pet trade, introduction of exotic species, infectious diseases, and climatic change. Unfortunately, almost none of these issues have been investigated for key populations in the Yucata´n Peninsula. For amphibians, studies exploring the presence of the chytrid fungus (Batracho- chytrium dendrobatidis) and the effects of climatic change are badly needed to understand the specific factors that negatively affect populations in this area. In general, conservation efforts for reptiles and amphibians in the Yucata´n Peninsula need to include environmental education, scientific investigation, and law enforce- ment and application. -

Reptiles of Ecuador: a Resource-Rich Online Portal, with Dynamic
Offcial journal website: Amphibian & Reptile Conservation amphibian-reptile-conservation.org 13(1) [General Section]: 209–229 (e178). Reptiles of Ecuador: a resource-rich online portal, with dynamic checklists and photographic guides 1Omar Torres-Carvajal, 2Gustavo Pazmiño-Otamendi, and 3David Salazar-Valenzuela 1,2Museo de Zoología, Escuela de Ciencias Biológicas, Pontifcia Universidad Católica del Ecuador, Avenida 12 de Octubre y Roca, Apartado 17- 01-2184, Quito, ECUADOR 3Centro de Investigación de la Biodiversidad y Cambio Climático (BioCamb) e Ingeniería en Biodiversidad y Recursos Genéticos, Facultad de Ciencias de Medio Ambiente, Universidad Tecnológica Indoamérica, Machala y Sabanilla EC170301, Quito, ECUADOR Abstract.—With 477 species of non-avian reptiles within an area of 283,561 km2, Ecuador has the highest density of reptile species richness among megadiverse countries in the world. This richness is represented by 35 species of turtles, fve crocodilians, and 437 squamates including three amphisbaenians, 197 lizards, and 237 snakes. Of these, 45 species are endemic to the Galápagos Islands and 111 are mainland endemics. The high rate of species descriptions during recent decades, along with frequent taxonomic changes, has prevented printed checklists and books from maintaining a reasonably updated record of the species of reptiles from Ecuador. Here we present Reptiles del Ecuador (http://bioweb.bio/faunaweb/reptiliaweb), a free, resource-rich online portal with updated information on Ecuadorian reptiles. This interactive portal includes encyclopedic information on all species, multimedia presentations, distribution maps, habitat suitability models, and dynamic PDF guides. We also include an updated checklist with information on distribution, endemism, and conservation status, as well as a photographic guide to the reptiles from Ecuador. -

Captive Wildlife Regulations, 2021, W-13.12 Reg 5
1 CAPTIVE WILDLIFE, 2021 W-13.12 REG 5 The Captive Wildlife Regulations, 2021 being Chapter W-13.12 Reg 5 (effective June 1, 2021). NOTE: This consolidation is not official. Amendments have been incorporated for convenience of reference and the original statutes and regulations should be consulted for all purposes of interpretation and application of the law. In order to preserve the integrity of the original statutes and regulations, errors that may have appeared are reproduced in this consolidation. 2 W-13.12 REG 5 CAPTIVE WILDLIFE, 2021 Table of Contents PART 1 PART 5 Preliminary Matters Zoo Licences and Travelling Zoo Licences 1 Title 38 Definition for Part 2 Definitions and interpretation 39 CAZA standards 3 Application 40 Requirements – zoo licence or travelling zoo licence PART 2 41 Breeding and release Designations, Prohibitions and Licences PART 6 4 Captive wildlife – designations Wildlife Rehabilitation Licences 5 Prohibition – holding unlisted species in captivity 42 Definitions for Part 6 Prohibition – holding restricted species in captivity 43 Standards for wildlife rehabilitation 7 Captive wildlife licences 44 No property acquired in wildlife held for 8 Licence not required rehabilitation 9 Application for captive wildlife licence 45 Requirements – wildlife rehabilitation licence 10 Renewal 46 Restrictions – wildlife not to be rehabilitated 11 Issuance or renewal of licence on terms and conditions 47 Wildlife rehabilitation practices 12 Licence or renewal term PART 7 Scientific Research Licences 13 Amendment, suspension, -
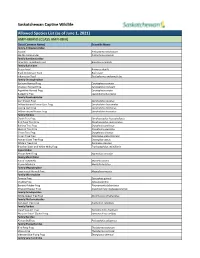
Captive Wildlife Allowed List
Saskatchewan Captive Wildlife Allowed Species List (as of June 1, 2021) AMPHIBIANS (CLASS AMPHIBIA) Class (Common Name) Scientific Name Family Ambystomatidae Axolotl Ambystoma mexicanum Marble Salamander Ambystoma opacum Family Bombinatoridae Oriental Fire-Bellied Toad Bombina orientalis Family Bufonidae Green Toad Anaxyrus debilis Black Indonesian Toad Bufo asper Indonesian Toad Duttaphrynus melanostictus Family Ceratophryidae Surinam Horned Frog Ceratophrys cornuta Chacoan Horned Frog Ceratophrys cranwelli Argentine Horned Frog Ceratophrys ornata Budgett’s Frog Lepidobatrachus laevis Family Dendrobatidae Dart Poison Frog Dendrobates auratus Yellow-banded Poison Dart Frog Dendrobates leucomelas Dyeing Dart Frog Dendrobates tinctorius Yellow-striped Poison Frog Dendrobates truncatus Family Hylidae Clown Tree Frog Dendropsophus leucophyllatus Bird Poop Tree Frog Dendropsophus marmoratus Barking Tree Frog Dryophytes gratiosus Squirrel Tree Frog Dryophytes squirellus Green Tree Frog Dryophytes cinereus Cuban Tree Frog Osteopilus septentrionalis Haitian Giant Tree Frog Osteopilus vastus White’s Tree Frog Ranoidea caerulea Brazilian Black and White Milky Frog Trachycephalus resinifictrix Hyperoliidae African Reed Frog Hyperolius concolor Family Mantellidae Baron’s Mantella Mantella baroni Brown Mantella Mantella betsileo Family Megophryidae Long-nosed Horned Frog Megophrys nasuta Family Microhylidae Tomato Frog Dyscophus guineti Chubby Frog Kaloula pulchra Banded Rubber Frog Phrynomantis bifasciatus Emerald Hopper Frog Scaphiophryne madagascariensis