Multiple Roles of Phosphoinositide-Specific Phospholipase C Isozymes
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

FAK Nuclear Export Signal Sequences
FEBS Letters 582 (2008) 2402–2406 FAK nuclear export signal sequences Valeria Ossovskayaa,1, Ssang-Taek Limb, Nobuyuki Otac, David D. Schlaepferb, Dusko Ilicc,d,* a Department of Anatomy, University of California San Francisco, San Francisco, CA, USA b Department of Reproductive Medicine, University of California, San Diego, CA, USA c A-cube Inc., Burlingame, CA, USA d Department of Cell and Tissue Biology, University of California San Francisco, San Francisco, CA, USA Received 13 April 2008; revised 28 May 2008; accepted 1 June 2008 Available online 10 June 2008 Edited by Varda Rotter accumulation. R177/R178A mutations also prevented FERM Abstract Ubiquitously expressed focal adhesion kinase (FAK), a critical component in transducing signals from sites of cell con- nuclear localization [12]. tacts with extracellular matrix, was named after its typical local- Since it is found in both cytoplasm and nucleus, FAK obvi- ization in focal adhesions. A nuclear localization of FAK has ously has to have a mechanism that enables nucleocytoplasmic been also reported and its scaffolding role in nucleus and require- shuttling. Leucine-rich nuclear export signal (NES) sequences ment for p53 ubiquitination were only recently described. often mediate protein export from the nucleus to the cyto- Whereas FAK nuclear localization signal (NLS) was found in plasm [13–16]. The first NES were identified in human immu- F2 lobe of FERM domain, nuclear export signal (NES) nodeficiency virus, type I-coded Rev protein [17] and protein sequences have not been yet determined. Here we demonstrate kinase A inhibitor of cAMP-dependent protein kinase [18]. that FAK has two NES sequences, NES1 in F1 lobe of FERM NES sequences consist of 4–5 hydrophobic residues within a domain and NES2 in kinase domain. -

Bioinformatic Analysis of Structure and Function of LIM Domains of Human Zyxin Family Proteins
International Journal of Molecular Sciences Article Bioinformatic Analysis of Structure and Function of LIM Domains of Human Zyxin Family Proteins M. Quadir Siddiqui 1,† , Maulik D. Badmalia 1,† and Trushar R. Patel 1,2,3,* 1 Alberta RNA Research and Training Institute, Department of Chemistry and Biochemistry, University of Lethbridge, 4401 University Drive, Lethbridge, AB T1K 3M4, Canada; [email protected] (M.Q.S.); [email protected] (M.D.B.) 2 Department of Microbiology, Immunology and Infectious Disease, Cumming School of Medicine, University of Calgary, 3330 Hospital Drive, Calgary, AB T2N 4N1, Canada 3 Li Ka Shing Institute of Virology, University of Alberta, Edmonton, AB T6G 2E1, Canada * Correspondence: [email protected] † These authors contributed equally to the work. Abstract: Members of the human Zyxin family are LIM domain-containing proteins that perform critical cellular functions and are indispensable for cellular integrity. Despite their importance, not much is known about their structure, functions, interactions and dynamics. To provide insights into these, we used a set of in-silico tools and databases and analyzed their amino acid sequence, phylogeny, post-translational modifications, structure-dynamics, molecular interactions, and func- tions. Our analysis revealed that zyxin members are ohnologs. Presence of a conserved nuclear export signal composed of LxxLxL/LxxxLxL consensus sequence, as well as a possible nuclear localization signal, suggesting that Zyxin family members may have nuclear and cytoplasmic roles. The molecular modeling and structural analysis indicated that Zyxin family LIM domains share Citation: Siddiqui, M.Q.; Badmalia, similarities with transcriptional regulators and have positively charged electrostatic patches, which M.D.; Patel, T.R. -

Nucleocytoplasmic Transport in Apoptosis
Nucleocytoplasmic transport in apoptosis E Ferrando-May*,1 Introduction 1 Molecular Toxicology Group, Faculty of Biology, University of Konstanz, PO The separation of the nucleus and the cytoplasm is the Box X911, 78457 Konstanz, Germany defining feature of eukaryotic cells and is achieved by the * Corresponding author: E Ferrando-May; Tel: þ 49 7531 884054; nuclear envelope, a double-membrane system of highly Fax: þ 49 7531 884033; E-mail: [email protected] selective permeability. Interchange of material between these two compartments occurs through dedicated transport chan- nels perforating the nuclear envelope, the nuclear pore complexes (NPCs). These are elaborate supramolecular structures consisting of about 30 different proteins, most of Abstract which are termed nucleoporins (Nups). The composition and structure of the NPC have been analysed in detail by a The apoptotic demolition of the nucleus is accomplished by combination of proteomics and electron microscopy ap- diverse proapoptotic factors, most of which are activated in proaches both in yeast and vertebrates, leading to a refined the cytoplasm and gain access to the nucleoplasm during the view of its molecular architecture. Essentially, the NPC is cell death process. The nucleus is also the main target for composed of three substructures of eight-fold rotational genotoxic insult, a potent apoptotic trigger. Signals generated symmetry: the cytoplasmic fibrils, the central framework, in the nucleus by DNA damage have to propagate to all and the nuclear basket (Figure 1). In the central framework, cellular compartments to ensure the coordinated execution of Nups form distinct subcomplexes which are arranged cell demise. The nucleocytoplasmic shuttling of signalling symmetrically with respect to the plane of the nuclear and execution factors is thus an integral part of the apoptotic envelope and enclose the central pore channel. -
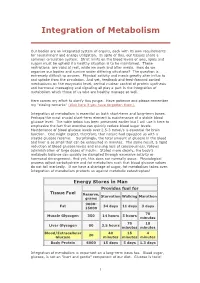
Integration of Metabolism
Integration of Metabolism Our bodies are an integrated system of organs, each with its own requirements for nourishment and energy utilization. In spite of this, our tissues share a common circulation system. Strict limits on the blood levels of ions, lipids and sugars must be upheld if a healthy situation is to be maintained. These restrictions are valid at rest, while we work and after meals. How do we organize our bodies and survive under differing situations? The question is extremely difficult to answer. Physical activity and meals greatly alter influx to and uptake from the circulation. And yet, feedback and feed-forward control mechanisms on the enzymatic level, central nuclear control of protein synthesis and hormonal messaging and signaling all play a part in the integration of metabolism which those of us who are healthy manage so well. Here comes my effort to clarify this jungle. Have patience and please remember my "closing remarks" (click here if you have forgotten them). Integration of metabolism is essential on both short-term and long-term bases. Perhaps the most crucial short-term element is maintenance of a stable blood glucose level. The table below has been presented earlier but I will use it here to emphasize the fact that exercise can quickly reduce blood sugar levels. Maintenance of blood glucose levels over 2.5-3 mmol/s is essential for brain function. One might expect, therefore, that nature had equipped us with a sizable glucose reserve. Surprisingly, the total amount of glucose in the blood and liver is so small that can be exhausted in minutes. -

Diagnostic Value of Serum Enzymes-A Review on Laboratory Investigations
Review Article ISSN 2250-0480 VOL 5/ ISSUE 4/OCT 2015 DIAGNOSTIC VALUE OF SERUM ENZYMES-A REVIEW ON LABORATORY INVESTIGATIONS. 1VIDYA SAGAR, M.SC., 2DR. VANDANA BERRY, MD AND DR.ROHIT J. CHAUDHARY, MD 1Vice Principal, Institute of Allied Health Sciences, Christian Medical College, Ludhiana 2Professor & Ex-Head of Microbiology Christian Medical College, Ludhiana 3Assistant Professor Department of Biochemistry Christian Medical College, Ludhiana ABSTRACT Enzymes are produced intracellularly, and released into the plasma and body fluids, where their activities can be measured by their abilities to accelerate the particular chemical reactions they catalyze. But different serum enzymes are raised when different tissues are damaged. So serum enzyme determination can be used both to detect cellular damage and to suggest its location in situ. Some of the biochemical markers such as alanine aminotransferase, aspartate aminotransferase, alkaline phasphatase, gamma glutamyl transferase, nucleotidase, ceruloplasmin, alpha fetoprotein, amylase, lipase, creatine phosphokinase and lactate dehydrogenase are mentioned to evaluate diseases of liver, pancreas, skeletal muscle, bone, etc. Such enzyme test may assist the physician in diagnosis and treatment. KEYWORDS: Liver Function tests, Serum Amylase, Lipase, CPK and LDH. INTRODUCTION mitochondrial AST is seen in extensive tissue necrosis during myocardial infarction and also in chronic Liver diseases like liver tissue degeneration DIAGNOSTIC SERUM ENZYME and necrosis². But lesser amounts are found in Enzymes are very helpful in the diagnosis of brain, pancreas and lung. Although GPT is plentiful cardiac, hepatic, pancreatic, muscular, skeltal and in the liver and occurs only in the small amount in malignant disorders. Serum for all enzyme tests the other tissues. -

GC-MS Metabolic Profile and -Glucosidase-, -Amylase-, Lipase-, and Acetylcholinesterase-Inhibitory Activities of Eight Peach
molecules Article GC-MS Metabolic Profile and α-Glucosidase-, α-Amylase-, Lipase-, and Acetylcholinesterase-Inhibitory Activities of Eight Peach Varieties Dasha Mihaylova 1,* , Ivelina Desseva 2,* , Aneta Popova 3 , Ivayla Dincheva 4 , Radka Vrancheva 2 , Anna Lante 5 and Albert Krastanov 1 1 Department of Biotechnology, Technological Faculty, University of Food Technologies, 4002 Plovdiv, Bulgaria; [email protected] 2 Department of Analytical Chemistry and Physical Chemistry, Technological Faculty, University of Food Technologies, 4002 Plovdiv, Bulgaria; [email protected] 3 Department of Catering and Tourism, Economics Faculty, University of Food Technologies, 4002 Plovdiv, Bulgaria; [email protected] 4 AgroBioInstitute, Agricultural Academy, 8 Dr. Tsankov Blvd., 1164 Sofia, Bulgaria; [email protected] 5 Department of Agronomy, Food, Natural Resources, Animals, and Environment—DAFNAE, Agripolis, University of Padova, 35020 Legnaro, Italy; [email protected] * Correspondence: [email protected] (D.M.); [email protected] (I.D.) Abstract: The inhibition of certain digestive enzymes by target food matrices represents a new approach in the treatment of socially significant diseases. Proving the ability of fruits to inhibit such enzymes can support the inclusion of specific varieties in the daily diets of patients with diabetes, obesity, Alzheimer’s disease, etc., providing them with much more than just valuable Citation: Mihaylova, D.; Desseva, I.; micro- and macromolecules. The current study aimed atidentifying and comparing the GC-MS Popova, A.; Dincheva, I.; Vrancheva, metabolic profiles of eight peach varieties (“Filina”, “Ufo 4, “Gergana”, “Laskava”, “July Lady”, R.; Lante, A.; Krastanov, A. GC-MS “Flat Queen”, “Evmolpiya”, and “Morsiani 90”) grown in Bulgaria (local and introduced) and to Metabolic Profile and α-Glucosidase-, evaluate the inhibitory potential of their extracts towards α-glucosidase, α-amylase, lipase, and α-Amylase-, Lipase-, and acetylcholinesterase. -

The Role of Phospholipase D (Pld) and Grb2 in Chemotaxis
THE ROLE OF PHOSPHOLIPASE D (PLD) AND GRB2 IN CHEMOTAXIS A thesis submitted in partial fulfillment of the requirements for the degree of Master of Science By KATIE J. KNAPEK B.S., Indiana University of Pennyslvania, 2006 2008 Wright State University WRIGHT STATE UNIVERSITY SCHOOL OF GRADUATE STUDIES December 19, 2008 I HEREBY RECOMMEND THAT THE THESIS PREPARED UNDER MY SUPERVISON BY Katie J. Knapek ENTITLED The Role of Phospholipase D (PLD) and Grb2 in Chemotaxis BE ACCEPTED IN PARTIAL FULFILLMENT OF THE REQUIREMENTS FOR THE DEGREE OF Master of Science. _____________________________ Julian Gomez-Cambronero, Ph. D. Thesis Director _____________________________ Barbara Hull, Ph. D. Program Director Committee on Final Examination _______________________ Julian Gomez-Cambronero, Ph. D. _______________________ Nancy Bigley, Ph. D. _______________________ Mill Miller, Ph. D. ________________________ Joseph F. Thomas Jr., Ph. D. Dean, School of Graduate Studies ABSTRACT Knapek, Katie J. M.S., Department of Biological Sciences, Microbiology and Immunology Program, Wright State University, 2008. The Role of Phospholipase D (PLD) and Grb2 in Chemotaxis. Phospholipase D (PLD) is an enzyme that hydrolyzes phosphatidylcholine yielding choline and phosphatidic acid. PLD is activated by mitogens (lead to cell division) and motogens (leading to cell migration). PLD is known to contribute to cellular proliferation and deregulated expression of PLD has been implicated in several human cancers. PLD has been found to play a role in leukocyte chemotaxis and adhesion as studied through the formation of chemokine gradients. We have established a model of cell migration comprising three cell lines: macrophages RAW 264.7 and LR-5 (for innate defense), and fibroblast COS-7 cells (for wound healing). -

Pancreatic Amylase and Lipase Plasma Concentrations Are
Diabetes Care Volume 38, May 2015 e71 Pancreatic Amylase and Lipase Plasma David P. Sonne,1 Tina Vilsbøll,1 and Filip K. Knop1,2 Concentrations Are Unaffected by Increments in Endogenous GLP-1 Levels Following Liquid Meal Tests Diabetes Care 2015;38:e71–e72 | DOI: 10.2337/dc14-2751 Recent findings have suggested that amylase and lipase in plasma following patients with type 2 diabetes (P , incretin-based therapies promote pan- the ingestion of oral glucose and three 0.05), whereas lipase concentrations creatic inflammation and possibly cell isocaloric and isovolemic liquid mealsd were similar. Neither of the enzymes proliferation within the endocrine and all of which exerted normal endogenous increased following nutrient ingestion, exocrine pancreas (1). However, these GLP-1 secretion (4)din patients with suggesting that postprandial elevations studies have been met with substan- type 2 diabetes and matched control of endogenous GLP-1 (two- to three- tive criticism based on technical and subjects. fold) cannot trigger enzyme release methodical issues (2). Nevertheless, Detailed description of the experimen- from the human pancreas, at least not incretin-based therapies seem to result tal procedures and subjects was provided acutely. in small increases (within normal range) previously (4). In short, pancreas-specific These results suggest that the obser- in plasma concentrations of amylase amylase and lipase concentrations were vation of elevated plasma amylase and andlipaseinpatientsreceivingthese measured in plasma from 15 patients lipase -

Exploring Machine Learning Methods for Nuclear Export Sequence Identification
Exploring Machine Learning Methods for Nuclear Export Sequence Identification A Major Qualifying Project Report Submitted to the Faculty Of the WORCESTER POLYTECHNIC INSTITUTE In partial fulfillment of the requirements for the Degree of Bachelor of Science ______________________________ Erin Conneilly Advisors: ______________________________ Professor Destin Heilman (CBC) ______________________________ Professor Rodica Neamtu (CS) Abstract The goal of this project is to design and implement a user-friendly machine learning tool that can be applied to the classification of polypeptides to find functional nuclear export sequences (NESs). This tool incorporates an API that takes advantage of support vector machines and can be expanded to include other models. Because NESs have been found to have consistent structure, structural data is incorporated into the model to increase confidence. This report is accompanied by a manual that instructs users on how to use the tool. 1 Table of Contents Abstract 1 Table of Contents 2 Table of Figures 3 Table of Tables 3 Acknowledgements 4 Introduction 5 Project Motivation 5 Problem Statement 5 Background 6 Section 1: Intracellular Transport 6 Section 2: Nuclear Export Sequence Structure 8 Section 3: Existing Prediction Methods 9 Section 4: Machine Learning Techniques 9 4.1 Recurrent Neural Networks (RNNs) 10 4.2 Nuclear Export Sequences as Time Series 10 4.3 Autoregressive Integrated Moving Average Model (ARIMA) 11 4.4 One-Class Support Vector Machines (OC SVM) 11 4.5 Scikit-Learn 13 Tool Overview and -

Functional Role of Pentose Phosphate Pathway and Glutamine in Cancer Cell Metabolism Ibrahim Halil Polat
Functional role of pentose phosphate pathway and glutamine in cancer cell metabolism Ibrahim Halil Polat To cite this version: Ibrahim Halil Polat. Functional role of pentose phosphate pathway and glutamine in cancer cell metabolism. Human health and pathology. Universitat de Barcelona, 2016. English. NNT : 2016GREAS031. tel-01722703 HAL Id: tel-01722703 https://tel.archives-ouvertes.fr/tel-01722703 Submitted on 5 Mar 2018 HAL is a multi-disciplinary open access L’archive ouverte pluridisciplinaire HAL, est archive for the deposit and dissemination of sci- destinée au dépôt et à la diffusion de documents entific research documents, whether they are pub- scientifiques de niveau recherche, publiés ou non, lished or not. The documents may come from émanant des établissements d’enseignement et de teaching and research institutions in France or recherche français ou étrangers, des laboratoires abroad, or from public or private research centers. publics ou privés. THÈSE Pour obtenir le grade de DOCTEUR DE LA COMMUNAUTÉ UNIVERSITÉ GRENOBLE ALPES préparée dans le cadre d’une cotutelle entre la Communauté Université Grenoble Alpes et Université de Barcelona Spécialité : BIS-Biotechnologie, instrumentation, signal et imagerie pour la biologie, la médecine et l’environnement Arrêté ministériel : 25 mai 2016 Présentée par « Ibrahim Halil Polat » Thèse dirigée par « Philippe Sabatier » codirigée par « Marta Cascante » préparée au sein des Laboratoires « TIMC-IMAG » et « BQI » dans les Écoles Doctorales « EDISCE » et « Ecole Doctorale de l’Université de Barcelone » Rôle fonctionnel des pentoses phosphates et glutamine dans le métabolisme des cellules cancéreuses Thèse soutenue publiquement le « 4 novembre 2016 », devant le jury composé de : M. Santiago Imperial Rodenas Professeur à l’Université de Barcelone (Président) Mme Loranne Agius Professeur à l’Université de Newcastle Upon Tyne (Rapporteur) M. -

Subcellular Localization of Crma: Identification of a Novel Leucine-Rich
Biochem. J. (2003) 373, 251–259 (Printed in Great Britain) 251 Subcellular localization of CrmA: identification of a novel leucine-rich nuclear export signal conserved in anti-apoptotic serpins Jose A. RODRIGUEZ1,Simone W. SPAN, Frank A. E. KRUYT and Giuseppe GIACCONE Department of Medical Oncology, VU University Medical Center, HV1081 Amsterdam, The Netherlands The cowpox virus-encoded anti-apoptotic protein cytokine flanking the NES contribute to the CRM1-dependent nuclear response modifier A (CrmA) is a member of the serpin family export of CrmA. Although YFP-tagged CrmA is primarily located that specifically inhibits the cellular proteins caspase 1, caspase in the cytoplasm, shifting its localization to be predominantly 8andgranzyme B. In this study, we have used Flag- and yellow nuclear by fusion of a heterologous nuclear localization signal fluorescent protein (YFP)-tagged versions of CrmA to investigate did not impair its ability to prevent Fas-induced apoptosis. We the mechanisms that regulate its subcellular localization. We show propose that nucleocytoplasmic shuttling would allow CrmA to that CrmA can actively enter and exit the nucleus and we dem- efficiently target cellular pro-apoptotic proteins not only in the onstrate the role of the nuclear export receptor CRM1 in this cytoplasm, but also in the nucleus, and thus to carry out its anti- shuttling process. CrmA contains a novel leucine-rich nuclear apoptotic function in both compartments. export signal (NES) that is functionally conserved in the anti- apoptotic cellular serpin PI-9. Besides this leucine-rich export signal, additional sequences mapping to a 103-amino-acid region Keywords: apoptosis, nuclear transport, CRM1. -

Unless Otherwise Noted, This Material Is Made Available
Author(s): Aken Desai, Michael Mathis, 2008 License: Unless otherwise noted, this material is made available under the terms of the Creative Commons Attribution – Share Alike 3.0 License: http://creativecommons.org/licenses/by-sa/3.0/ We have reviewed this material in accordance with U.S. Copyright Law and have tried to maximize your ability to use, share, and adapt it. Copyright holders of content included in this material should contact [email protected] with any quesCons, correcCons, or clarificaon regarding the use of content. For more informaon about how to cite these materials visit hHp://open.umich.edu/educaon/about/terms-of-use. Student works are presented as is and may be an interpretaon of faculty members’ lectures or assignments. These student works are not a product of faculty members. Faculty do not guarantee the accuracy of student work nor endorse them in any way. Any medical informaon in this material is intended to inform and educate and is not a tool for self-diagnosis or a replacement for medical evaluaon, advice, diagnosis or treatment by a healthcare professional. Please speak to your physician if you have quesCons about your medical condiCon. Viewer discreon is advised: Some medical content is graphic and may not be suitable for all viewers. Fatty Acids ctd. Tuesday, January 15, 2008 10:00 AM 14. What is the role of pentose pathway? Why is it important in fatty acid biosynthesis? What regulates the pentose pathway? How are pentoses formed without the oxidation of glucose‐6‐phosphate? a. Role is to provide NADPH and convert glucose into 5 carbon sugars for other pathways b.