Lower Paleogene Crocodilians from Silveirinha, Portugal
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

JVP 26(3) September 2006—ABSTRACTS
Neoceti Symposium, Saturday 8:45 acid-prepared osteolepiforms Medoevia and Gogonasus has offered strong support for BODY SIZE AND CRYPTIC TROPHIC SEPARATION OF GENERALIZED Jarvik’s interpretation, but Eusthenopteron itself has not been reexamined in detail. PIERCE-FEEDING CETACEANS: THE ROLE OF FEEDING DIVERSITY DUR- Uncertainty has persisted about the relationship between the large endoskeletal “fenestra ING THE RISE OF THE NEOCETI endochoanalis” and the apparently much smaller choana, and about the occlusion of upper ADAM, Peter, Univ. of California, Los Angeles, Los Angeles, CA; JETT, Kristin, Univ. of and lower jaw fangs relative to the choana. California, Davis, Davis, CA; OLSON, Joshua, Univ. of California, Los Angeles, Los A CT scan investigation of a large skull of Eusthenopteron, carried out in collaboration Angeles, CA with University of Texas and Parc de Miguasha, offers an opportunity to image and digital- Marine mammals with homodont dentition and relatively little specialization of the feeding ly “dissect” a complete three-dimensional snout region. We find that a choana is indeed apparatus are often categorized as generalist eaters of squid and fish. However, analyses of present, somewhat narrower but otherwise similar to that described by Jarvik. It does not many modern ecosystems reveal the importance of body size in determining trophic parti- receive the anterior coronoid fang, which bites mesial to the edge of the dermopalatine and tioning and diversity among predators. We established relationships between body sizes of is received by a pit in that bone. The fenestra endochoanalis is partly floored by the vomer extant cetaceans and their prey in order to infer prey size and potential trophic separation of and the dermopalatine, restricting the choana to the lateral part of the fenestra. -

The World at the Time of Messel: Conference Volume
T. Lehmann & S.F.K. Schaal (eds) The World at the Time of Messel - Conference Volume Time at the The World The World at the Time of Messel: Puzzles in Palaeobiology, Palaeoenvironment and the History of Early Primates 22nd International Senckenberg Conference 2011 Frankfurt am Main, 15th - 19th November 2011 ISBN 978-3-929907-86-5 Conference Volume SENCKENBERG Gesellschaft für Naturforschung THOMAS LEHMANN & STEPHAN F.K. SCHAAL (eds) The World at the Time of Messel: Puzzles in Palaeobiology, Palaeoenvironment, and the History of Early Primates 22nd International Senckenberg Conference Frankfurt am Main, 15th – 19th November 2011 Conference Volume Senckenberg Gesellschaft für Naturforschung IMPRINT The World at the Time of Messel: Puzzles in Palaeobiology, Palaeoenvironment, and the History of Early Primates 22nd International Senckenberg Conference 15th – 19th November 2011, Frankfurt am Main, Germany Conference Volume Publisher PROF. DR. DR. H.C. VOLKER MOSBRUGGER Senckenberg Gesellschaft für Naturforschung Senckenberganlage 25, 60325 Frankfurt am Main, Germany Editors DR. THOMAS LEHMANN & DR. STEPHAN F.K. SCHAAL Senckenberg Research Institute and Natural History Museum Frankfurt Senckenberganlage 25, 60325 Frankfurt am Main, Germany [email protected]; [email protected] Language editors JOSEPH E.B. HOGAN & DR. KRISTER T. SMITH Layout JULIANE EBERHARDT & ANIKA VOGEL Cover Illustration EVELINE JUNQUEIRA Print Rhein-Main-Geschäftsdrucke, Hofheim-Wallau, Germany Citation LEHMANN, T. & SCHAAL, S.F.K. (eds) (2011). The World at the Time of Messel: Puzzles in Palaeobiology, Palaeoenvironment, and the History of Early Primates. 22nd International Senckenberg Conference. 15th – 19th November 2011, Frankfurt am Main. Conference Volume. Senckenberg Gesellschaft für Naturforschung, Frankfurt am Main. pp. 203. -

Artiodactyla and Perissodactyla (Mammalia) from the Early-Middle Eocene Kuldana Formation of Kohat (Pakistan)
CO"uTK1BL 11015 FKOLI IHt \lC5tLL1 OF I' ALEO\ IOLOG1 THE UNIVERSITY OF IVICHIGAN VOI 77 Lo 10 p 717-37.1 October 33 1987 ARTIODACTYLA AND PERISSODACTYLA (MAMMALIA) FROM THE EARLY-MIDDLE EOCENE KULDANA FORMATION OF KOHAT (PAKISTAN) BY J. G. M. THEWISSEN. P. D. GINGERICH and D. E. RUSSELL MUSEUM OF PALEONTOLOGY THE UNIVERSITY OF MICHIGAN ANN ARBOR CONTRIBUTIONS FROM THE MUSEUM OF PALEONTOLOGY Charles B. Beck, Director Jennifer A. Kitchell, Editor This series of contributions from the Museum of Paleontology is a medium for publication of papers based chiefly on collections in the Museum. When the number of pages issued is sufficient to make a volume, a title page and a table of contents will be sent to libraries on the mailing list, and to individuals upon request. A list of the separate issues may also be obtained by request. Correspond- ence should be directed to the Museum of Paleontology, The University of Michigan, Ann Arbor, Michigan 48109. VOLS. II-XXVII. Parts of volumes may be obtained if available. Price lists are available upon inquiry. I ARTIODACTI L .-I A\D PERISSODACTYL4 (kl.iihlhlAL1A) FROM THE EARLY-h1IDDLE EOCEUE KCLD..I\4 FORMATIO\ OF KOHAT (PAKISTAY) J. G. M. THEWISSEN. P. D. GINGERICH AND D. E. RUSSELL Ah.strcict.-Chorlakki. yielding approximately 400 specimens (mostly isolated teeth and bone fragments). is one of four major early-to-middle Eocene niammal localities on the Indo-Pakistan subcontinent. On the basis of ung~~latesclescribed in this paper we consider the Chorlakki fauna to be younger than that from Barbora Banda. -

Gyraulus Laevis in Nederland 87
Kuijper: Gyraulus laevis in Nederland 87 Gyraulus laevis (Mollusca: Planorbidae) in Nederland door W.J. Kuijper Enkele recente waarnemingen van een van onze zeldzaamste planorbiden waren de aanleiding tot het samenstellen van een over- laevis zicht van alle van Nederland bekende vondsten van Gyraulus (Alder, 1838). Tot voor kort waren slechts enkele vindplaatsen van dit dier bekend (Janssen & De Vogel, 1965: 75). Hierbij komt het betrof feit dat het in een aantal gevallen slechts een enkel exemplaar en de soort niet meer teruggevonden kon worden. Het volgende geeft een chronologisch overzicht van de waarnemingen van Gyraulus laevis in Nederland. Voor zover beschikbaar zijn diverse gegevens van de vindplaatsen vermeld. WAARNEMINGEN 1. Koudekerke (bij Middelburg), buitenplaats Vijvervreugd, ± 1890. Fraai vers materiaal: 69 exemplaren in de collectie Schep- de man (Zoölogisch Museum, Amsterdam). Later niet meer vermeld, buitenplaats bestaat niet meer (Kuiper, 1944: 6). Dit materiaalwerd verzameld door Dr. IJ. Keijzer en is vermeld in: Verslag over 1885-1893 van het Zeeuwsch Genootschap der Wetenschappen, blz. 88 (volgens Mevr. Dr. W.S.S. van der Feen - van Benthem Jut- ting). Verder is materiaal van deze vindplaats in de collecties van het Zeeuwsch Museum te Middelburg (2 ex.) en het Natuurhistorisch Museum te Enschede (6 ex.) aanwezig. 2. Warmond (bij Leiden), in de Leede, januari 1916 (Van Ben- them Jutting, 1933: 179). Dit materiaal werd verzameld door P.P. de Koning; in het Rijksmuseum van Natuurlijke Historie te Leiden bevinden zich vier schelpen van deze vindplaats, waarvan twee een doorsnede van ca. 4 mm bereiken. 3. 't Zand (bij Roodeschool), Oostpolder, 1927 (Van Benthem Jutting, 1947: 66). -
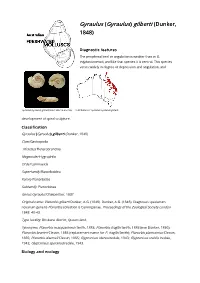
Gyraulus) Gilberti (Dunker, 1848
Gyraulus (Gyraulus) gilberti (Dunker, 1848) Diagnostic features The peripheral keel or angulation is weaker than in G. edgbastonensis, and like that species it is central. This species varies widely in degree of depression and angulation, and Gyraulus (Gyraulus) gilberti (adult size 4.8-5.5 mm) Distribution of Gyraulus (Gyraulus) gilberti. development of spiral sculpture. Classification Gyraulus (Gyraulus) gilberti (Dunker, 1848) Class Gastropoda I nfraclass Heterobranchia Megaorder Hygrophila Order Lymnaeida Superfamily Planorboidea Family Planorbidae Subfamily: Planorbinae Genus Gyraulus Charpentier, 1837 Original name: Planorbis gilberti Dunker, A.G. (1848). Dunker, A.G. (1848). Diagnoses specierum novarum generis Planorbis collection is Cumingianae. Proceedings of the Zoological Society London 1848: 40-43. Type locality: Brisbane district, Queensland. Synonyms: Planorbis macquariensis Smith, 1883; Planorbis fragilis Smith, 1883 (non Dunker, 1850); Planorbis brazieri Clessin, 1885 (replacement name for P. fragilis Smith); Planorbis planissimus Clessin, 1885; Planorbis daemeli Clessin, 1885; Glyptanisus idenusredale, 1943; Glyptanisus stabilis redale, 1943; Glyptanisus speranusredale, 1943. Biology and ecology This species lives in water weeds and other vegetation in ponds, billabongs, swamps and sluggish streams and rivers in tropical and subtropical eastern Australia. Feeds on detritus. Egg mass presumably a jelly strip containing small eggs. Development direct. Brown (2001) described the anatomy of this species. This species is an intermediate host for the stomach fluke Orthocoelium streptocoelium (Boray, 1982; Beesley et al., 1998). Distribution This species occurs throughout eastern Australia, from Cape York to northern New South Wales. Notes G. isingi and/or G.waterhousei may possibly be conspecific with this species (Brown, 2001). Further reading Beesley, P. L., Ross, G. J. -

Anisus Vorticulus (Troschel 1834) (Gastropoda: Planorbidae) in Northeast Germany
JOURNAL OF CONCHOLOGY (2013), VOL.41, NO.3 389 SOME ECOLOGICAL PECULIARITIES OF ANISUS VORTICULUS (TROSCHEL 1834) (GASTROPODA: PLANORBIDAE) IN NORTHEAST GERMANY MICHAEL L. ZETTLER Leibniz Institute for Baltic Sea Research Warnemünde, Seestr. 15, D-18119 Rostock, Germany Abstract During the EU Habitats Directive monitoring between 2008 and 2010 the ecological requirements of the gastropod species Anisus vorticulus (Troschel 1834) were investigated in 24 different waterbodies of northeast Germany. 117 sampling units were analyzed quantitatively. 45 of these units contained living individuals of the target species in abundances between 4 and 616 individuals m-2. More than 25.300 living individuals of accompanying freshwater mollusc species and about 9.400 empty shells were counted and determined to the species level. Altogether 47 species were identified. The benefit of enhanced knowledge on the ecological requirements was gained due to the wide range and high number of sampled habitats with both obviously convenient and inconvenient living conditions for A. vorticulus. In northeast Germany the amphibian zones of sheltered mesotrophic lake shores, swampy (lime) fens and peat holes which are sun exposed and have populations of any Chara species belong to the optimal, continuously and densely colonized biotopes. The cluster analysis emphasized that A. vorticulus was associated with a typical species composition, which can be named as “Anisus-vorticulus-community”. In compliance with that both the frequency of combined occurrence of species and their similarity in relative abundance are important. The following species belong to the “Anisus-vorticulus-community” in northeast Germany: Pisidium obtusale, Pisidium milium, Pisidium pseudosphaerium, Bithynia leachii, Stagnicola palustris, Valvata cristata, Bathyomphalus contortus, Bithynia tentaculata, Anisus vortex, Hippeutis complanatus, Gyraulus crista, Physa fontinalis, Segmentina nitida and Anisus vorticulus. -

Attachment J Assessment of Existing Paleontologic Data Along with Field Survey Results for the Jonah Field
Attachment J Assessment of Existing Paleontologic Data Along with Field Survey Results for the Jonah Field June 12, 2007 ABSTRACT This is compilation of a technical analysis of existing paleontological data and a limited, selective paleontological field survey of the geologic bedrock formations that will be impacted on Federal lands by construction associated with energy development in the Jonah Field, Sublette County, Wyoming. The field survey was done on approximately 20% of the field, primarily where good bedrock was exposed or where there were existing, debris piles from recent construction. Some potentially rich areas were inaccessible due to biological restrictions. Heavily vegetated areas were not examined. All locality data are compiled in the separate confidential appendix D. Uinta Paleontological Associates Inc. was contracted to do this work through EnCana Oil & Gas Inc. In addition BP and Ultra Resources are partners in this project as they also have holdings in the Jonah Field. For this project, we reviewed a variety of geologic maps for the area (approximately 47 sections); none of maps have a scale better than 1:100,000. The Wyoming 1:500,000 geology map (Love and Christiansen, 1985) reveals two Eocene geologic formations with four members mapped within or near the Jonah Field (Wasatch – Alkali Creek and Main Body; Green River – Laney and Wilkins Peak members). In addition, Winterfeld’s 1997 paleontology report for the proposed Jonah Field II Project was reviewed carefully. After considerable review of the literature and museum data, it became obvious that the portion of the mapped Alkali Creek Member in the Jonah Field is probably misinterpreted. -

Gyraulus) Meridionalis (Brazier, 1875
Gyraulus (Gyraulus) meridionalis (Brazier, 1875) Diagnostic features The periphery is subangled to distinctly angled, with angulation in middle or slightly towards upper surface (viewed with aperture on right). Some fine axial and spiral sculpture. The shell can reach Gyraulus (Gyraulus) meridionalis (adult size up to 5 mm) (lectotype) Distribution of Gyraulus (Gyraulus) meridionalis. about 5 mm in maximum diameter. Classification Gyraulus (Gyraulus) meridionalis (Brazier, 1875) Class Gastropoda I nfraclass Heterobranchia Megaorder Hygrophila Order Lymnaeida Superfamily Planorboidea Family Planorbidae Subfamily: Planorbinae Genus Gyraulus Charpentier, 1837 Original name: Planorbis meridionalis Brazier, 1875. Brazier, J. (1875). Descriptions of eight species of Australian and Tasmanian land and freshwater shells. Proceedings of the Linnean Society of New South Wales 1: 17-20. Type locality: Circular Head, Tasmania. Synonyms: Planorbis tasmanicus Tenison Woods, 1876; Planorbis brazieri Clessin, 1885 (in part). Biology and ecology This species lives on aquatic vegetation and stones in temperate ponds, streams and rivers of south- eastern Australia. Feeds on detritus. Egg mass presumably a jelly strip containing small eggs. Development direct. Brown (1998 & 2001) described the anatomy of this species. Distribution Tasmania, southern Victoria and south-eastern South Australia. Notes Brown (2001) considered that the distinct reproductive anatomy of G. meridionalis potentially justify a distinct subgeneric status for this species. Further reading Brown, D. S. (1981). Observations on the Planorbidae from Australia and New Guinea. Journal of the Malacological Society of Australia 5: 67-80. Brown, D. S. (1998). Freshwater snails of the genus Gyraulus (Gastropoda: Planorbidae) in Australia: the taxa of Tasmania. Molluscan Research 19: 105-154. Brown, D. S. (2001). -

Gyraulus) Waterhousei (Clessin, 1885
Gyraulus (Gyraulus) waterhousei (Clessin, 1885) Diagnostic features This keeled species has a depressed shell and is variably spirally sculptured. Classification Gyraulus (Gyraulus) waterhousei (adult size up to 4.3 mm) Gyraulus (Gyraulus) waterhousei (adult size up to Distribution of Gyraulus (Gyraulus) waterhousei. 4.3 mm) Gyraulus (Gyraulus) waterhousei (Clessin, 1885) Class Gastropoda I nfraclass Heterobranchia Megaorder Hygrophila Order Lymnaeida Superfamily Planorboidea Family Planorbidae Subfamily: Planorbinae Genus Gyraulus Charpentier, 1837 Original name: Planorbis waterhousei Clessin, 1885. Clessin, S. (1885). Die Familie der Limnaeiden enthaltend die Genera Planorbis, Limnaeus, Physa und Amphipeplea. Systematisches Conchylien- Cabinet 1(17): 223-310, plates 40-50. Type locality: Clarence River, NSW. Synonyms: Glyptanisus metaurus redale,1943; Glyptanisus ordessus redale, 1943. Biology and ecology This species lives on aquatic vegetation in ponds, billabongs, swamps, streams and rivers. Feeds on detritus. Egg mass presumably a jelly strip containing small eggs. Development direct. nhabits coastal areas. Distribution Eastern New South Wales and possibly southern Queensland. Notes Brown (2001) considers that this species may possibly be a synonym of G. gilberti. Further reading Brown, D. S. (1981). Observations on the Planorbidae from Australia and New Guinea. Journal of the Malacological Society of Australia 5: 67-80. Brown, D. S. (1998). Freshwater snails of the genus Gyraulus (Gastropoda: Planorbidae) in Australia: the taxa of Tasmania. Molluscan Research 19: 105-154. Brown, D. S. (2001). Freshwater snails of the genus Gyraulus (Planorbidae) in Australia: taxa of the mainland. Molluscan Research 21: 17- 107. Hubendick, B. (1955). Phylogeny of the Planorbidae. Transactions of the Zoological Society of London 28: 453-542. Shea, M. -

Rapid and Early Post-Flood Mammalian Diversification Videncede in the Green River Formation
The Proceedings of the International Conference on Creationism Volume 6 Print Reference: Pages 449-457 Article 36 2008 Rapid and Early Post-Flood Mammalian Diversification videncedE in the Green River Formation John H. Whitmore Cedarville University Kurt P. Wise Southern Baptist Theological Seminary Follow this and additional works at: https://digitalcommons.cedarville.edu/icc_proceedings DigitalCommons@Cedarville provides a publication platform for fully open access journals, which means that all articles are available on the Internet to all users immediately upon publication. However, the opinions and sentiments expressed by the authors of articles published in our journals do not necessarily indicate the endorsement or reflect the views of DigitalCommons@Cedarville, the Centennial Library, or Cedarville University and its employees. The authors are solely responsible for the content of their work. Please address questions to [email protected]. Browse the contents of this volume of The Proceedings of the International Conference on Creationism. Recommended Citation Whitmore, John H. and Wise, Kurt P. (2008) "Rapid and Early Post-Flood Mammalian Diversification Evidenced in the Green River Formation," The Proceedings of the International Conference on Creationism: Vol. 6 , Article 36. Available at: https://digitalcommons.cedarville.edu/icc_proceedings/vol6/iss1/36 In A. A. Snelling (Ed.) (2008). Proceedings of the Sixth International Conference on Creationism (pp. 449–457). Pittsburgh, PA: Creation Science Fellowship and Dallas, TX: Institute for Creation Research. Rapid and Early Post-Flood Mammalian Diversification Evidenced in the Green River Formation John H. Whitmore, Ph.D., Cedarville University, 251 N. Main Street, Cedarville, OH 45314 Kurt P. Wise, Ph.D., Southern Baptist Theological Seminary, 2825 Lexington Road. -

Contributions in BIOLOGY and GEOLOGY
MILWAUKEE PUBLIC MUSEUM Contributions In BIOLOGY and GEOLOGY ~umber 56 March 21, 1984 Paleontology and Geology of the Bridger Formation, ,outhern Green River Basin, Southwestern Wyoming. Part 7. Survey of Bridgerian Artiodactyla, including description of a skull and partial skeleton of Antiacodon pygmaeus. Robert M. West REVIEWERS FOR THIS PUBLICATION: Robert J. Emry, National Museum of Natural History Peter Sheehan, Milwaukee Public Museum John A. Wilson, Balcones Research Center, Austin, Texas ISBN 0-89326-099-1 © 1984Milwaukee Public Museum Published by the order of the Board of Trustees Paleontology and Geology of the Bridger Formation, Southern Green River Basin, Southwestern Wyoming. Part 7. Survey of Bridgerian Artiodactyla, including description of a skull and partial skeleton of Antiacodon pygmaeus. Robert M. West Carnegie Museum of Natural History Pittsburgh, PA 15213 ACKNOWLEDGMENTS I am grateful to Robert J. Emry, U.S. National Museum of Natural His- tory, Washington, D.C.; Mary R. Dawson, Carnegie Museum of Natural History, Pittsburgh, PA; Richard H. Tedford, American Museum of Natural History, New York, NY; Leo J. Hickey and Mary Anne Turner, Yale Peabody Museum of Natural History, New Haven, CT; Donald Baird, Princeton Uni- versity, Princeton, NJ; and William D. Turnbull, Field Museum of Natural History, Chicago, IL, for access to and loan of specimens in their respective charges. Margery C. Coombs, University of Massachusetts, Amherst, MA, provided pre-publication information on her study of artiodactyl ear regions, and Kenneth D. Rose, The Johns Hopkins University, commented on the postcranial anatomy of Antiacodon and allowed me to examine material of Diacodexis. The drawings of Antiacodon were prepared by Susan D. -

Download Full Article in PDF Format
Relations et position systématique du genre Cuisitherium Sudre et al., 1983, le plus dérivé des artiodactyles de l’Éocène inférieur d’Europe Jean SUDRE Laboratoire de Paléontologie des Vertébrés E.P.H.E., ISEM Université de Montpellier 2, case courrier 064, Place Eugène Bataillon, F-34095 Montpellier cedex 05 (France) [email protected] Gérard LECOMTE 5 Rue Guiscard, F-60400 Grisolles (France) Sudre J. & Lecomte G. 2000. — Relations et position systématique du genre Cuisitherium Sudre et al., 1983, le plus dérivé des artiodactyles de l’Éocène inférieur d’Europe. Geodiversitas 22 (3) : 415-432. RÉSUMÉ La description d’un maxillaire de Cuisitherium lydekkeri découvert sur le gi- sement yprésien de Monthelon a permis de préciser les caractères dentaires de l’espèce ainsi que les relations et le statut systématique du genre Cuisitherium. On établit que Cuisitherium est probablement directement apparenté au genre monospécifique Lophiobunodon connu dans les niveaux élevés de l’Éocène moyen (La Livinière, MP15). La relation Cuisitherium- Lophiobunodon montre que l’acquisition des molaires supérieures dilambdo- dontes (ectolophe en W et pli de la centrocrête) a dû se réaliser rapidement au MOTS CLÉS cours de l’Éocène moyen sans que cela entraîne des modifications majeures Mammifère, dans la morphologie des molaires inférieures. Cette relation conduit à ratta- artiodactyle, cher Cuisitherium aux Choeropotamidae, conformément à l’opinion de Éocène, évolution, Gentry & Hooker (1988), cette famille incluant d’après ces auteurs des systématique. formes rapportées antérieurement aux Haplobunodontidae. GEODIVERSITAS • 2000 • 22 (3) © Publications Scientifiques du Muséum national d’Histoire naturelle, Paris. www.mnhn.fr/publication/ 415 Sudre J.