The First Systematic Description of Cambrian Fossils from Myanmar
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-
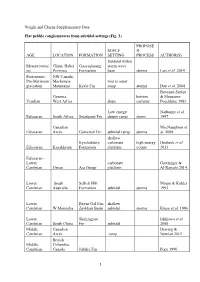
Wright and Cherns Supplementary Data Flat
Wright and Cherns Supplementary Data Flat pebble conglomerates from subtidal settings (Fig. 2) PROPOSE SHELF D AGE LOCATION FORMATION SETTING PROCESS AUTHOR(S) Subtidal within Mesoproteroz China, Hebei Gaoyuzhuang storm wave oic Province Formation base storms Luo et al. 2014 Proterozoic, NW Canada, Pre-Marinoan Mackenzie mid to outer glaciation Mountains Keele Fm ramp storms Day et al. 2004 Bertrand-Sarfati Gourma, bottom & Moussine- Vendian West Africa slope currents' Pouchkine 1983 Low energy Narbonne et al. Ediacaran South Africa Swartpunt Fm deeper ramp storm 1997 Canadian MacNaughton et Ediacaran Arctic Gametrail Fm subtidal ramp storms al. 2008 shallow Kyrshabakty carbonate high energy Heubeck et al. Ediacaran Kazakhstan Formation platform events 2013 Ediacaran - Lower carbonate Grotzinger & Cambrian Oman Ara Group platform Al-Rawahi 2014 Lower South Sellick Hill Mount & Kidder Cambrian Australia Formation subtidal storms 1993 Lower Bayan Gol Fm, shallow Cambrian W Mongolia Zavkhan Basin subtidal storms Kruse et al. 1996 Lower Shuijingtuo Ishikawa et al. Cambrian South China Fm subtidal 2008 Middle Canadian Dewing & Cambrian Arctic ramp Nowlan 2012 British Middle Columbia, Cambrian Canada Jubilee Fm Pope 1990 1 Middle Pratt & Cambrian Argentina La Laja Fm subtidal shelf tsunamis Bordonaro 2007 low energy Middle shallow Cambrian Australia Ranken Lst subtidal storms Kruse 1996 Middle Wyoming, Upper Gros Cambrian USA Ventre Shale Csonka 2009 middle upper Middle W Utah, upper Wheeler, carbonate belt - Cambrian USA Marjum fms subtidal shelf Robison 1964 Middle- Upper Supratidal to Cambrian NW China subtidal fpc storms Liang et al. 1993 Ust’- Brus,Labaz, Middle Orakta, Cambrian - Kulyumbe, Lower Ujgur and Iltyk Kouchinsky et Ordovician Siberia fms al. -

And Ordovician (Sardic) Felsic Magmatic Events in South-Western Europe: Underplating of Hot Mafic Magmas Linked to the Opening of the Rheic Ocean
Solid Earth, 11, 2377–2409, 2020 https://doi.org/10.5194/se-11-2377-2020 © Author(s) 2020. This work is distributed under the Creative Commons Attribution 4.0 License. Comparative geochemical study on Furongian–earliest Ordovician (Toledanian) and Ordovician (Sardic) felsic magmatic events in south-western Europe: underplating of hot mafic magmas linked to the opening of the Rheic Ocean J. Javier Álvaro1, Teresa Sánchez-García2, Claudia Puddu3, Josep Maria Casas4, Alejandro Díez-Montes5, Montserrat Liesa6, and Giacomo Oggiano7 1Instituto de Geociencias (CSIC-UCM), Dr. Severo Ochoa 7, 28040 Madrid, Spain 2Instituto Geológico y Minero de España, Ríos Rosas 23, 28003 Madrid, Spain 3Dpt. Ciencias de la Tierra, Universidad de Zaragoza, 50009 Zaragoza, Spain 4Dpt. de Dinàmica de la Terra i de l’Oceà, Universitat de Barcelona, Martí Franquès s/n, 08028 Barcelona, Spain 5Instituto Geológico y Minero de España, Plaza de la Constitución 1, 37001 Salamanca, Spain 6Dpt. de Mineralogia, Petrologia i Geologia aplicada, Universitat de Barcelona, Martí Franquès s/n, 08028 Barcelona, Spain 7Dipartimento di Scienze della Natura e del Territorio, 07100 Sassari, Italy Correspondence: J. Javier Álvaro ([email protected]) Received: 1 April 2020 – Discussion started: 20 April 2020 Revised: 14 October 2020 – Accepted: 19 October 2020 – Published: 11 December 2020 Abstract. A geochemical comparison of early Palaeo- neither metamorphism nor penetrative deformation; on the zoic felsic magmatic episodes throughout the south- contrary, their unconformities are associated with foliation- western European margin of Gondwana is made and in- free open folds subsequently affected by the Variscan defor- cludes (i) Furongian–Early Ordovician (Toledanian) activ- mation. -

Cambrian Cephalopods
BULLETIN 40 Cambrian Cephalopods BY ROUSSEAU H. FLOWER 1954 STATE BUREAU OF MINES AND MINERAL RESOURCES NEW MEXICO INSTITUTE OF MINING & TECHNOLOGY CAMPUS STATION SOCORRO, NEW MEXICO NEW MEXICO INSTITUTE OF MINING & TECHNOLOGY E. J. Workman, President STATE BUREAU OF MINES AND MINERAL RESOURCES Eugene Callaghan, Director THE REGENTS MEMBERS Ex OFFICIO The Honorable Edwin L. Mechem ...................... Governor of New Mexico Tom Wiley ......................................... Superintendent of Public Instruction APPOINTED MEMBERS Robert W. Botts ...................................................................... Albuquerque Holm 0. Bursum, Jr. ....................................................................... Socorro Thomas M. Cramer ........................................................................ Carlsbad Frank C. DiLuzio ..................................................................... Los Alamos A. A. Kemnitz ................................................................................... Hobbs Contents Page ABSTRACT ...................................................................................................... 1 FOREWORD ................................................................................................... 2 ACKNOWLEDGMENTS ............................................................................. 3 PREVIOUS REPORTS OF CAMBRIAN CEPHALOPODS ................ 4 ADEQUATELY KNOWN CAMBRIAN CEPHALOPODS, with a revision of the Plectronoceratidae ..........................................................7 -

Appendix 3.Pdf
A Geoconservation perspective on the trace fossil record associated with the end – Ordovician mass extinction and glaciation in the Welsh Basin Item Type Thesis or dissertation Authors Nicholls, Keith H. Citation Nicholls, K. (2019). A Geoconservation perspective on the trace fossil record associated with the end – Ordovician mass extinction and glaciation in the Welsh Basin. (Doctoral dissertation). University of Chester, United Kingdom. Publisher University of Chester Rights Attribution-NonCommercial-NoDerivatives 4.0 International Download date 26/09/2021 02:37:15 Item License http://creativecommons.org/licenses/by-nc-nd/4.0/ Link to Item http://hdl.handle.net/10034/622234 International Chronostratigraphic Chart v2013/01 Erathem / Era System / Period Quaternary Neogene C e n o z o i c Paleogene Cretaceous M e s o z o i c Jurassic M e s o z o i c Jurassic Triassic Permian Carboniferous P a l Devonian e o z o i c P a l Devonian e o z o i c Silurian Ordovician s a n u a F y r Cambrian a n o i t u l o v E s ' i k s w o Ichnogeneric Diversity k p e 0 10 20 30 40 50 60 70 S 1 3 5 7 9 11 13 15 17 19 21 n 23 r e 25 d 27 o 29 M 31 33 35 37 39 T 41 43 i 45 47 m 49 e 51 53 55 57 59 61 63 65 67 69 71 73 75 77 79 81 83 85 87 89 91 93 Number of Ichnogenera (Treatise Part W) Ichnogeneric Diversity 0 10 20 30 40 50 60 70 1 3 5 7 9 11 13 15 17 19 21 n 23 r e 25 d 27 o 29 M 31 33 35 37 39 T 41 43 i 45 47 m 49 e 51 53 55 57 59 61 c i o 63 z 65 o e 67 a l 69 a 71 P 73 75 77 79 81 83 n 85 a i r 87 b 89 m 91 a 93 C Number of Ichnogenera (Treatise Part W) -

The Late Cambrian SPICE Carbon Isotope Excursion
CORE Metadata, citation and similar papers at core.ac.uk Provided by NERC Open Research Archive The Furongian (late Cambrian) Steptoean Positive Carbon Isotope Excursion (SPICE) in Avalonia Mark A. Woods1*, Philip R. Wilby1, Melanie J. Leng2, Adrian W. A. Rushton1,3 & Mark Williams1,4 1: British Geological Survey, Keyworth, Nottingham, NG12 5GG, UK 2: NERC Isotope Geosciences Laboratory, British Geological Survey, Keyworth, Nottingham, NG12 5GG, UK 3: Department of Palaeontology, The Natural History Museum, Cromwell Road, South Kensington, London, SW7 5DB, UK 4: Department of Geology, University of Leicester, Leicester LE1 7RH, UK *Corresponding author Mark Woods (email: [email protected]) The Steptoean Positive Carbon Isotope Excursion (SPICE) of earliest late Cambrian (Furongian) age is identified in England. The excursion is found within a ca 145 m thick siliciclastic succession within the middle and higher part of the Outwoods Shale Formation of Warwickshire, and reaches a 13 maximum δ Corg amplitude of 4.1‰ at values of –25.6‰. Biostratigraphical data show that the excursion occupies the greater part of the Olenus trilobite biozone, an equivalent of the Glyptagnostus reticulatus biozone that marks the base of the Furongian and coeval base of the Steptoean in North America. The amplitude of the excursion approaches that recorded in limestone- dominated Laurentian successions, and is greater than that recently documented for organic-rich mudstones of palaeocontinental Baltica in southern Sweden. A minor positive excursion above the SPICE may equate with a similar excursion recognised in Siberia. The SPICE in the Outwoods Shale Formation seems closely linked to the widely recognised early Furongian eustatic sea level rise. -

PROGRAMME ABSTRACTS AGM Papers
The Palaeontological Association 63rd Annual Meeting 15th–21st December 2019 University of Valencia, Spain PROGRAMME ABSTRACTS AGM papers Palaeontological Association 6 ANNUAL MEETING ANNUAL MEETING Palaeontological Association 1 The Palaeontological Association 63rd Annual Meeting 15th–21st December 2019 University of Valencia The programme and abstracts for the 63rd Annual Meeting of the Palaeontological Association are provided after the following information and summary of the meeting. An easy-to-navigate pocket guide to the Meeting is also available to delegates. Venue The Annual Meeting will take place in the faculties of Philosophy and Philology on the Blasco Ibañez Campus of the University of Valencia. The Symposium will take place in the Salon Actos Manuel Sanchis Guarner in the Faculty of Philology. The main meeting will take place in this and a nearby lecture theatre (Salon Actos, Faculty of Philosophy). There is a Metro stop just a few metres from the campus that connects with the centre of the city in 5-10 minutes (Line 3-Facultats). Alternatively, the campus is a 20-25 minute walk from the ‘old town’. Registration Registration will be possible before and during the Symposium at the entrance to the Salon Actos in the Faculty of Philosophy. During the main meeting the registration desk will continue to be available in the Faculty of Philosophy. Oral Presentations All speakers (apart from the symposium speakers) have been allocated 15 minutes. It is therefore expected that you prepare to speak for no more than 12 minutes to allow time for questions and switching between presenters. We have a number of parallel sessions in nearby lecture theatres so timing will be especially important. -

001-012 Primeras Páginas
PUBLICACIONES DEL INSTITUTO GEOLÓGICO Y MINERO DE ESPAÑA Serie: CUADERNOS DEL MUSEO GEOMINERO. Nº 9 ADVANCES IN TRILOBITE RESEARCH ADVANCES IN TRILOBITE RESEARCH IN ADVANCES ADVANCES IN TRILOBITE RESEARCH IN ADVANCES planeta tierra Editors: I. Rábano, R. Gozalo and Ciencias de la Tierra para la Sociedad D. García-Bellido 9 788478 407590 MINISTERIO MINISTERIO DE CIENCIA DE CIENCIA E INNOVACIÓN E INNOVACIÓN ADVANCES IN TRILOBITE RESEARCH Editors: I. Rábano, R. Gozalo and D. García-Bellido Instituto Geológico y Minero de España Madrid, 2008 Serie: CUADERNOS DEL MUSEO GEOMINERO, Nº 9 INTERNATIONAL TRILOBITE CONFERENCE (4. 2008. Toledo) Advances in trilobite research: Fourth International Trilobite Conference, Toledo, June,16-24, 2008 / I. Rábano, R. Gozalo and D. García-Bellido, eds.- Madrid: Instituto Geológico y Minero de España, 2008. 448 pgs; ils; 24 cm .- (Cuadernos del Museo Geominero; 9) ISBN 978-84-7840-759-0 1. Fauna trilobites. 2. Congreso. I. Instituto Geológico y Minero de España, ed. II. Rábano,I., ed. III Gozalo, R., ed. IV. García-Bellido, D., ed. 562 All rights reserved. No part of this publication may be reproduced or transmitted in any form or by any means, electronic or mechanical, including photocopy, recording, or any information storage and retrieval system now known or to be invented, without permission in writing from the publisher. References to this volume: It is suggested that either of the following alternatives should be used for future bibliographic references to the whole or part of this volume: Rábano, I., Gozalo, R. and García-Bellido, D. (eds.) 2008. Advances in trilobite research. Cuadernos del Museo Geominero, 9. -

Lee-Riding-2018.Pdf
Earth-Science Reviews 181 (2018) 98–121 Contents lists available at ScienceDirect Earth-Science Reviews journal homepage: www.elsevier.com/locate/earscirev Marine oxygenation, lithistid sponges, and the early history of Paleozoic T skeletal reefs ⁎ Jeong-Hyun Leea, , Robert Ridingb a Department of Geology and Earth Environmental Sciences, Chungnam National University, Daejeon 34134, Republic of Korea b Department of Earth and Planetary Sciences, University of Tennessee, Knoxville, TN 37996, USA ARTICLE INFO ABSTRACT Keywords: Microbial carbonates were major components of early Paleozoic reefs until coral-stromatoporoid-bryozoan reefs Cambrian appeared in the mid-Ordovician. Microbial reefs were augmented by archaeocyath sponges for ~15 Myr in the Reef gap early Cambrian, by lithistid sponges for the remaining ~25 Myr of the Cambrian, and then by lithistid, calathiid Dysoxia and pulchrilaminid sponges for the first ~25 Myr of the Ordovician. The factors responsible for mid–late Hypoxia Cambrian microbial-lithistid sponge reef dominance remain unclear. Although oxygen increase appears to have Lithistid sponge-microbial reef significantly contributed to the early Cambrian ‘Explosion’ of marine animal life, it was followed by a prolonged period dominated by ‘greenhouse’ conditions, as sea-level rose and CO2 increased. The mid–late Cambrian was unusually warm, and these elevated temperatures can be expected to have lowered oxygen solubility, and to have promoted widespread thermal stratification resulting in marine dysoxia and hypoxia. Greenhouse condi- tions would also have stimulated carbonate platform development, locally further limiting shallow-water cir- culation. Low marine oxygenation has been linked to episodic extinctions of phytoplankton, trilobites and other metazoans during the mid–late Cambrian. -

Abstract Volume
https://doi.org/10.3301/ABSGI.2019.04 Milano, 2-5 July 2019 ABSTRACT BOOK a cura della Società Geologica Italiana 3rd International Congress on Stratigraphy GENERAL CHAIRS Marco Balini, Università di Milano, Italy Elisabetta Erba, Università di Milano, Italy - past President Società Geologica Italiana 2015-2017 SCIENTIFIC COMMITTEE Adele Bertini, Peter Brack, William Cavazza, Mauro Coltorti, Piero Di Stefano, Annalisa Ferretti, Stanley C. Finney, Fabio Florindo, Fabrizio Galluzzo, Piero Gianolla, David A.T. Harper, Martin J. Head, Thijs van Kolfschoten, Maria Marino, Simonetta Monechi, Giovanni Monegato, Maria Rose Petrizzo, Claudia Principe, Isabella Raffi, Lorenzo Rook ORGANIZING COMMITTEE The Organizing Committee is composed by members of the Department of Earth Sciences “Ardito Desio” and of the Società Geologica Italiana Lucia Angiolini, Cinzia Bottini, Bernardo Carmina, Domenico Cosentino, Fabrizio Felletti, Daniela Germani, Fabio M. Petti, Alessandro Zuccari FIELD TRIP COMMITTEE Fabrizio Berra, Mattia Marini, Maria Letizia Pampaloni, Marcello Tropeano ABSTRACT BOOK EDITORS Fabio M. Petti, Giulia Innamorati, Bernardo Carmina, Daniela Germani Papers, data, figures, maps and any other material published are covered by the copyright own by the Società Geologica Italiana. DISCLAIMER: The Società Geologica Italiana, the Editors are not responsible for the ideas, opinions, and contents of the papers published; the authors of each paper are responsible for the ideas opinions and con- tents published. La Società Geologica Italiana, i curatori scientifici non sono responsabili delle opinioni espresse e delle affermazioni pubblicate negli articoli: l’autore/i è/sono il/i solo/i responsabile/i. ST3.2 Cambrian stratigraphy, events and geochronology Conveners and Chairpersons Per Ahlberg (Lund University, Sweden) Loren E. -

Ediacaran and Cambrian Stratigraphy in Estonia: an Updated Review
Estonian Journal of Earth Sciences, 2017, 66, 3, 152–160 https://doi.org/10.3176/earth.2017.12 Ediacaran and Cambrian stratigraphy in Estonia: an updated review Tõnu Meidla Department of Geology, Institute of Ecology and Earth Sciences, Faculty of Science and Technology, University of Tartu, Ravila 14a, 50411 Tartu, Estonia; [email protected] Received 18 December 2015, accepted 18 May 2017, available online 6 July 2017 Abstract. Previous late Precambrian and Cambrian correlation charts of Estonia, summarizing the regional stratigraphic nomenclature of the 20th century, date back to 1997. The main aim of this review is updating these charts based on recent advances in the global Precambrian and Cambrian stratigraphy and new data from regions adjacent to Estonia. The term ‘Ediacaran’ is introduced for the latest Precambrian succession in Estonia to replace the formerly used ‘Vendian’. Correlation with the dated sections in adjacent areas suggests that only the latest 7–10 Ma of the Ediacaran is represented in the Estonian succession. The gap between the Ediacaran and Cambrian may be rather substantial. The global fourfold subdivision of the Cambrian System is introduced for Estonia. The lower boundary of Series 2 is drawn at the base of the Sõru Formation and the base of Series 3 slightly above the former lower boundary of the ‘Middle Cambrian’ in the Baltic region, marked by a gap in the Estonian succession. The base of the Furongian is located near the base of the Petseri Formation. Key words: Ediacaran, Cambrian, correlation chart, biozonation, regional stratigraphy, Estonia, East European Craton. INTRODUCTION The latest stratigraphic chart of the Cambrian System in Estonia (Mens & Pirrus 1997b, p. -

GSSP) of the Drumian Stage (Cambrian) in the Drum Mountains, Utah, USA
Articles 8585 by Loren E. Babcock1, Richard A. Robison2, Margaret N. Rees3, Shanchi Peng4, and Matthew R. Saltzman1 The Global boundary Stratotype Section and Point (GSSP) of the Drumian Stage (Cambrian) in the Drum Mountains, Utah, USA 1 School of Earth Sciences, The Ohio State University, 125 South Oval Mall, Columbus, OH 43210, USA. Email: [email protected] and [email protected] 2 Department of Geology, University of Kansas, Lawrence, KS 66045, USA. Email: [email protected] 3 Department of Geoscience, University of Nevada, Las Vegas, Las Vegas, NV 89145, USA. Email: [email protected] 4 State Key Laboratory of Palaeobiology and Stratigraphy, Nanjing Institute of Geology and Palaeontology, Chinese Academy of Sciences, 39 East Beijing Road, Nanjing 210008, China. Email: [email protected] The Global boundary Stratotype Section and Point correlated with precision through all major Cambrian regions. (GSSP) for the base of the Drumian Stage (Cambrian Among the methods that should be considered in the selection of a GSSP (Remane et al., 1996), biostratigraphic, chemostratigraphic, Series 3) is defined at the base of a limestone (cal- paleogeographic, facies-relationship, and sequence-stratigraphic cisiltite) layer 62 m above the base of the Wheeler For- information is available (e.g., Randolph, 1973; White, 1973; McGee, mation in the Stratotype Ridge section, Drum Moun- 1978; Dommer, 1980; Grannis, 1982; Robison, 1982, 1999; Rowell et al. 1982; Rees 1986; Langenburg et al., 2002a, 2002b; Babcock et tains, Utah, USA. The GSSP level contains the lowest al., 2004; Zhu et al., 2006); that information is summarized here. occurrence of the cosmopolitan agnostoid trilobite Pty- Voting members of the International Subcommission on Cam- chagnostus atavus (base of the P. -

The Auxiliary Boundary Stratotype Section and Point (ASSP) of the Jiangshanian Stage (Cambrian: Furongian Series) in the Kyrshabakty Section
See discussions, stats, and author profiles for this publication at: https://www.researchgate.net/publication/261721078 The Auxiliary boundary Stratotype Section and Point (ASSP) of the Jiangshanian Stage (Cambrian: Furongian Series) in the Kyrshabakty section Article in Episodes · March 2014 DOI: 10.18814/epiiugs/2014/v37i1/005 CITATION READS 1 238 7 authors, including: Leonid E. Popov Michael G. Bassett National Museum Wales National Museum of Wales 296 PUBLICATIONS 3,956 CITATIONS 46 PUBLICATIONS 557 CITATIONS SEE PROFILE SEE PROFILE Some of the authors of this publication are also working on these related projects: Early Palaeozoic brachiopods and associated faunas of CAOB. View project Cambrian-Ordovician Boundary View project All content following this page was uploaded by Leonid E. Popov on 21 April 2014. The user has requested enhancement of the downloaded file. 41 by Gappar Kh. Ergaliev1, Vyacheslav (Slava) G. Zhemchuzhnikov2, Leonid E. Popov3, Michael G. Bassett3 and Farkhat G. Ergaliev4 The Auxiliary boundary Stratotype Section and Point (ASSP) of the Jiangshanian Stage (Cambrian: Furongian Series) in the Kyrshabakty section, Kazakhstan 1 K.I.Satpaev Institute of Geological Sciences, Kabanbai Batyra Street, 69-a, 050010 Almaty, Kazakhstan. E-mail: [email protected] 2 North Caspian Oil Development LLP, Almaty, Kazakhstan. E-mail: [email protected] 3 Department of Geology, National Museum of Wales, Cathays Park, Cardiff CF10 3NP, Wales, United Kingdom. E-mail: [email protected] 4 Dala Mining (TMM International), Koshek-Batyra Street, 5, 050010 Almaty Kazakhstan. E-mail: [email protected] The Kyrshabakty section in the Malyi Karatau Range, Union of Geological Sciences (IUGS). The GSSP for the stage, now southern Kazakhstan, has been approved as the Auxiliary known as the Jiangshanian (formerly provisional Stage 9) is located in the Duibian B section, Zhejiang, China (Peng et al., 2012).