The Effect of Calcium and Halide Ions on the Gramicidin a Molecular State
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Influence of Water on Conductivity of Aluminium Halide Solutions in Acetonitrile
Influence of water on conductivity of aluminium halide solutions in acetonitrile L. LUX Department of Chemistry, Faculty of Metallurgy, Technical University, CS-043 85 Košice Received 7 September 1981 Accepted for publication 8 February 1982 Equilibria of the substitution and association reactions of AICI3 and AlBr3 with acetonitrile were investigated by measuring the conductivity of solutions at varying concentrations of A1C13, AlBr3, and water in the acetonitrile solution. Both halides behave in acetonitrile in the absence of water as relatively strong electrolytes. It has been revealed that water displaces acetoni trile from the solvate sphere of cation of both halides. As for AlBr3, the bromide ion is also displaced from the coordination sphere of aluminium and replaced by a neutral water molecule, which simultaneously results in an increased number of ionized particles in solution. The hydrate A1X3 -6H20 was identified as final product which separates from the solution by adding water. Изучены равновесия реакций замещения и ассоциации А1С13 и А1Вг3 с ацетонитрилом посредством измерения электропроводности раствора в зависимости от концентрации А1С13, А1Вг3, как и в зависимости от концентрации воды в растворе ацетонитрила. Оба галогенида ведут себя в ацетонитриле как сравнительно сильные электролиты в отсутствии воды. Обнаружено, что вода вытесняет ацетонитрил из сольватной обо лочки катиона у обоих галогенидов. У А1Вг3 доходит и к вытеснению иона брома из координационной сферы алюминия и его замещению нейтраль ной молекулой воды, что одновременно приводит к увеличению числа ионизированных частиц в растворе. Конечным продуктом, выделенным из раствора при добавлении воды был идентифицированный гидрат А1Х3-6Н20. Acetonitrile is a typical aprotic solvent which is widely used in electrochemical research because of its profitable properties, i.e. -

Molecular Dynamics Simulation of Ion Mobility. 2. Alkali Metal and Halide Ions Using the SPC/E Model for Water at 25 °C†
+ + 1420 J. Phys. Chem. 1996, 100, 1420-1425 Molecular Dynamics Simulation of Ion Mobility. 2. Alkali Metal and Halide Ions Using the SPC/E Model for Water at 25 °C† Song Hi Lee Department of Chemistry, Kyungsung UniVersity, Pusan 608-736, Korea Jayendran C. Rasaiah* Department of Chemistry, UniVersity of Maine, Orono, Maine 04469 ReceiVed: October 16, 1995; In Final Form: NoVember 10, 1995X We present results of computer simulations of the mobilities of the alkali metal ions (Li+, Na+,K+, Rb+, and Cs+) and the halides (F-, Cl-, Br-, and I-) at 25 °C using the SPC/E model for water and ion-water parameters fitted to the binding energies of small clusters of ions. A simple truncation of the ion-water and water- water potentials was used, and the mobilities calculated from the mean square displacement and the velocity autocorrelation functions, respectively, were found to be in good agreement with each other. The calculations demonstrate, for the first time, cation and anion mobilities that fall on separate curves, as functions of ion size, with distinct maxima. This is in complete accord with experimental trends observed in water at 25 °C. The cation mobilities are also in better agreement with the measured values than the calculations done earlier (J. Chem. Phys. 1994, 101, 6964) using the TIP4P model. The mobilities of the halides calculated here for the SPC/E model are however slightly lower than the experimental results. The residence times of water in the hydration shells around an ion are found to decrease dramatically with its size. Stereoscopic pictures show that the structure of the solvent cage around an ion is qualitatively different for the larger ions, implicating both solvent dynamics and structure as important factors in explaining ion mobility in aqueous systems. -

Halides and Halogens. What Do I Need to Know? John Vivari, Nordson EFD
Halides and Halogens. What do I need to know? John Vivari, Nordson EFD Abstract With halogen-containing substances in the public eye due to scrutiny by the European Union and a variety of non- governmental organizations (NGOs) as possible additions to the list of substances banned from electronics, we at EFD have received numerous inquiries from customers asking how this subject will affect them and their processes. Having just overcome the hurdle of RoHS (Restriction of Hazardous Substances), they want to know what halogens and halides are, and what changes they should be prepared for if required to stop using them. Halide-free materials are not new. Some segments of the electronics industry have been sensitive to halides and their significance for decades. This paper will give the reader a working knowledge of halogens and halides. Armed with this education, the reader will be able to make informed decisions when required to use halogen-free materials, either because regulations dictate it or social pressure makes acceptance preferable to resistance. Key Words: halide, halogen, bromine, chlorine, flame retardant, RoHS What are halogens and halides? damage. Brominated flame retardant use is not limited to electronics. It is also in common usage in furniture, At their most basic level, halogens are the electronegative construction materials and textiles. elements in column 17 of the periodic table, including fluorine (F), chlorine, (Cl), bromine (Br), iodine (I) and Other sources of halogens in circuit boards include astatine (At). In electronics fiberglass sizing, epoxy curing agents and accelerators, applications, iodine and resin wetting and de-foaming agents, flux residues, and astatine are rarely if ever contamination from handling. -
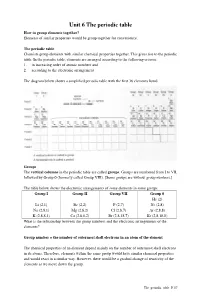
Unit 6 the Periodic Table How to Group Elements Together? Elements of Similar Properties Would Be Group Together for Convenience
Unit 6 The periodic table How to group elements together? Elements of similar properties would be group together for convenience. The periodic table Chemists group elements with similar chemical properties together. This gives rise to the periodic table. In the periodic table, elements are arranged according to the following criteria: 1. in increasing order of atomic numbers and 2. according to the electronic arrangement The diagram below shows a simplified periodic table with the first 36 elements listed. Groups The vertical columns in the periodic table are called groups . Groups are numbered from I to VII, followed by Group 0 (formerly called Group VIII). [Some groups are without group numbers.] The table below shows the electronic arrangements of some elements in some groups. Group I Group II Group VII Group 0 He (2) Li (2,1) Be (2,2) F (2,7) Ne (2,8) Na (2,8,1) Mg (2,8,2) Cl (2,8,7) Ar (2,8,8) K (2,8,8,1) Ca (2,8,8,2) Br (2,8,18,7) Kr (2,8,18,8) What is the relationship between the group numbers and the electronic arrangements of the elements? Group number = the number of outermost shell electrons in an atom of the element The chemical properties of an element depend mainly on the number of outermost shell electrons in its atoms. Therefore, elements within the same group would have similar chemical properties and would react in a similar way. However, there would be a gradual change of reactivity of the elements as we move down the group. -

Calcium Bromide Liquid: Safety and Handling Guide Geobrom® HG520
Calcium Bromide Liquid Safety and Handling Guide GeoBrom® HG520 Great Lakes Solutions: brominated derivative products for mercury control Contents Emergency Response Telephone Numbers 1 Responsible Care®– Doing the Right Thing 2 Introduction 3 GeoBrom® HG520 Product Description 5 Product Description Typical Product Quality Physical and Chemical Properties Environmental, Health, Safety and Regulatory Summary Regulatory Information and Training 9 Procedures for the Safe Handling of Calcium Bromide Liquid 11 Product Data Sheet Personal Protective Equipment (PPE) Safety and Handling Safety Considerations Environmental Considerations Spills Recommended Storage and Security Emergency Procedures 13 Emergency Equipment and Supplies Transportation Emergencies Fire First Aid 14 Equipment and Materials of Construction 16 Corrosion Studies Summary Shipping Containers and Transportation 17 Contact Information 19 Emergency Response Telephone Numbers for Bromine Country/ Emergency Response Provider and Region Emergency Number* CHEMTREC® United States +1-800-424-9300 or 1-703-527-3887 Carechem 24 Europe +44 (0) 1235 239 670 Carechem 24 India +91 1166 4114 05 National Registration Centre for Chemicals (NRCC) China 0532-8388-9090 *Refer to the Safety Data Sheet (SDS) for additional regional emergency response phone numbers. 1 | GLS GeoBrom® HG520 CaBr2 Liquid Safety and Handling Guide Responsible Care®– Doing the Right Thing Responsible Care® is the chemical Our Responsible Care® Policy industry’s health, safety, security, Chemtura considers ensuring the safety and health of the public, our employees, and environmental performance contractors, and customers and the protection of the environment now and improvement initiative. As an for future generations to be our core values. We are committed to meeting obligation of membership, all applicable laws, regulations, permits, and internal standards throughout Responsible Care® is the ethical the world. -

Zinc-Calcium Bromide Solution
TETRA Technologies, Inc. Material Safety Data Sheet Zinc-Calcium Bromide Solution 1. PRODUCT AND COMPANY IDENTIFICATION U.S. Toll Free (800) 424-9300 CHEMTREC (24-Hr Emergency Response): TETRA Technologies, Inc. International +1 703-527-3887 25025 I-45 North Poison Control: (800) 222-1222 The Woodlands, TX 77380 Non-emergency Customer Service: (281) 367-1983 Synonyms: Zinc-Calcium Bromide, Liquid Zinc-Calcium Bromide Chemical Family: Inorganic Salt Solution MSDS Creation Date: 08 Oct 96 MSDS Revision Date: 09 Apr 08 2. HAZARDS IDENTIFICATION NFPA Ratings: (Scale 0-4) Health: 2 Fire: 0 Reactivity: 0 WARNING: Irritant; Marine Pollutant; Explosion potential with mixtures containing potassium or sodium. Odorless, clear, colorless to amber liquid. May cause irritation to skin, eyes and respiratory system. Avoid breathing mists, EMERGENCY OVERVIEW: contact with eyes, skin and clothing. Harmful if swallowed. Do not ingest. Keep container tightly closed. Wash thoroughly after handling. Keep container tightly closed. Wash thoroughly after handling. Use only with adequate ventilation. Potential Health Effects Dermal sensitizer. Single, short exposure not likely to cause significant skin irritation. However, direct contact with solution may cause irritation, redness and Skin Contact: pain, possibly severe. Additional effects may include blistering, erythema, exfoliation, ulceration, necrosis, and scarring. May cause irritation of eyes with redness and pain and superficial injury. Additional Eye Contact: effects may include tearing, eye discharge, conjunctivitis, and blurred vision. Inhalation may cause irritation to mucous membranes and respiratory system. Inhalation: Additional effects may include coughing, labored breathing, and shortness of breath. Ingestion may cause burning sensation in mouth and throat, yellowing of the skin and eyes, nausea, vomiting, diarrhea, blood in urine, inability to urinate, low blood Ingestion: pressure, blood disorders, kidney damage, liver damage, convulsions and unconsciousness. -

Chemical Compatibility Chart for Plazit-Polygal PLAZGAL SAN Sheets
Chemical Compatibility Chart for Plazit-Polygal PLAZGAL SAN Sheets This table gives an indication of the chemical resistance of PLAZGAL SAN sheets to a range of common chemicals at 20ºC. PLAZGAL SAN sheets can be safely used with most chemicals and components such as alkalis and diluted acids, as well as with aqueous salt solutions. However, SAN has limited resistance to alcohols, aliphatic hydrocarbons, oils and fats, and is not resistant to concentrated mineral acids, aromatic and/ or halogenated hydrocarbons, esters, ethers, ketones. Important note: the chemical stability of PLAZGAL SAN sheets depends on many factors such as the concentration of the chemical agent, internal stresses in the sheets and exposure temperature, in certain situations the sheets can develop stress cracks when even when exposed to chemicals to which SAN is usually resistant. The resistance of PLAZGAL SAN sheets is indicated in the table below. Chemical Concentration Compliance Chemical Concentration Compliance Aqua Regia Dissolved Brake fluids (ATE) Affected Acetic acid To 50% Not affected Butter Not affected Acetic acid 100% Affected Butyl acetate Affected Acetone Affected Butyric acid Affected Acetylsalicylic (soln.) Not affected Cadmium (soln.) Not affected acid bromide Allyl alcohol Affected Caffeine (soln.) Not affected Almond, bitter, Not affected Calcium bromide (soln.) Not affected oil of Calcium chloride (soln.) Not affected Aluminium (soln.) Not affected Calcium (solid) Not affected chloride hypochlorite Aluminium (soln.) Not affected Calcium (soln.) -

The Elements.Pdf
A Periodic Table of the Elements at Los Alamos National Laboratory Los Alamos National Laboratory's Chemistry Division Presents Periodic Table of the Elements A Resource for Elementary, Middle School, and High School Students Click an element for more information: Group** Period 1 18 IA VIIIA 1A 8A 1 2 13 14 15 16 17 2 1 H IIA IIIA IVA VA VIAVIIA He 1.008 2A 3A 4A 5A 6A 7A 4.003 3 4 5 6 7 8 9 10 2 Li Be B C N O F Ne 6.941 9.012 10.81 12.01 14.01 16.00 19.00 20.18 11 12 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 3 Na Mg IIIB IVB VB VIB VIIB ------- VIII IB IIB Al Si P S Cl Ar 22.99 24.31 3B 4B 5B 6B 7B ------- 1B 2B 26.98 28.09 30.97 32.07 35.45 39.95 ------- 8 ------- 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 4 K Ca Sc Ti V Cr Mn Fe Co Ni Cu Zn Ga Ge As Se Br Kr 39.10 40.08 44.96 47.88 50.94 52.00 54.94 55.85 58.47 58.69 63.55 65.39 69.72 72.59 74.92 78.96 79.90 83.80 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52 53 54 5 Rb Sr Y Zr NbMo Tc Ru Rh PdAgCd In Sn Sb Te I Xe 85.47 87.62 88.91 91.22 92.91 95.94 (98) 101.1 102.9 106.4 107.9 112.4 114.8 118.7 121.8 127.6 126.9 131.3 55 56 57 72 73 74 75 76 77 78 79 80 81 82 83 84 85 86 6 Cs Ba La* Hf Ta W Re Os Ir Pt AuHg Tl Pb Bi Po At Rn 132.9 137.3 138.9 178.5 180.9 183.9 186.2 190.2 190.2 195.1 197.0 200.5 204.4 207.2 209.0 (210) (210) (222) 87 88 89 104 105 106 107 108 109 110 111 112 114 116 118 7 Fr Ra Ac~RfDb Sg Bh Hs Mt --- --- --- --- --- --- (223) (226) (227) (257) (260) (263) (262) (265) (266) () () () () () () http://pearl1.lanl.gov/periodic/ (1 of 3) [5/17/2001 4:06:20 PM] A Periodic Table of the Elements at Los Alamos National Laboratory 58 59 60 61 62 63 64 65 66 67 68 69 70 71 Lanthanide Series* Ce Pr NdPmSm Eu Gd TbDyHo Er TmYbLu 140.1 140.9 144.2 (147) 150.4 152.0 157.3 158.9 162.5 164.9 167.3 168.9 173.0 175.0 90 91 92 93 94 95 96 97 98 99 100 101 102 103 Actinide Series~ Th Pa U Np Pu AmCmBk Cf Es FmMdNo Lr 232.0 (231) (238) (237) (242) (243) (247) (247) (249) (254) (253) (256) (254) (257) ** Groups are noted by 3 notation conventions. -

“All in the Family” Properties of Halogens SCIENTIFIC Periodic Trends and the Properties of the Elements
“All in the Family” Properties of Halogens SCIENTIFIC Periodic Trends and the Properties of the Elements Introduction In every family there are similarities and differences. The same is true in chemical families as well. Elements that share similar chemical properties are arranged in vertical columns, called groups or families, in the modern periodic table. For example, the Group 17 elements, consisting of fluorine, chlorine, bromine, iodine, and astatine, are called the halogens. What are the similarities and differences in the chemical properties of the halogens and their compounds? Concepts • Halogens • Periodic table • Groups or families • Periodic trends Activity Overview The purpose of this demonstration is to explore the similarities and differences in the chemical properties of the halogens. The reactions of chlorine, bromine, and iodine with sodium chloride, sodium bromide, and sodium iodide will be inves- tigated in order to determine the periodic trend for the reactivity of the halogens. Chlorine, bromine, and iodine will be identified by their unique colors in water and hexane. Materials Bromine water, Br2 in H2O, 10 mL† Sodium thiosulfate solution, 4%, 500 mL§ Chlorine water, Cl2 in H2O, 10 mL† Water, distilled or deionized Iodine solution, I2, 0.04 M, 10 mL Bottles (with caps), square glass, 30-mL, 6 Hexanes, C6H14, 60 mL Graduated cylinders, 10- and 25-mL Sodium bromide solution, NaBr, 0.1 M, 10 mL Marking pencil or pen Sodium chloride solution, NaCl, 0.1 M, 10 mL Periodic table Sodium iodide solution, NaI, 0.1 M, 10 mL †Prepare fresh within one day of use—see the Preparation of Solutions section. -

THE PHYSICAL CHEMISTRY of METALS in THEIR MOLTEN HALIDES THESIS Presented for the Degree of DOCTOR of PHILOSOPHY in the UNIVERSI
THE PHYSICAL CHEMISTRY OF METALS IN THEIR MOLTEN HALIDES THESIS presented for the degree of DOCTOR OF PHILOSOPHY in the UNIVERSITY OF LONDON By Lars-Ingvar Staffansson, Bergsingenjor, (Stockholm). London, December 1959, ABSTRACT The phase diagrams of the systems calcium- calcium chloride, calcium-calcium bromide and calcium- calcium iodide have been established by solubility measurements and differential thermal analysis. Each of the systems was found to have a eutectic a mono- tectic and a large miscibility gap. The eutectics are 3.3, 3.2 and 6.8 mole % calcium respectively, and the corresponding temperatures are 763, 728 and 760°C. The monotectics are at 99.5, 99.6 and 99.7 mole % calcium respectively and the corresponding temperatures are 826, 828 and 831°C. The consolute points are at 1338, 1335 and 1380°C and the corresponding compositions are 62, 64 and 74 mole % calcium. Discrepancies between this and previous work are mainly attributed to composition changes of the phases during quenching. A method has been developed that overcomes this difficulty. The depression of the freezing point of strontium by strontium chloride has also been determined. From the results of this work a model for the mutal solubility of metalS and their molten halides has been suggested. CONTENTS Page CHAPTER 1: INTRODUCTION 1 1.1 General Introduction 1 1.2 Relevant previous work on metal-molten salts. 4 1.2.1 General 4 1.2.2 Alkaline earth metal-metal halide systems. 12 CHAPTER 2: EXPERIMENTAL 24 2.1 Programme 24 2.2 Preparation and purity of materials 25 2.2.1 Calcium 25 2.2.2 Strontium 26 2.2.3 Calcium Chloride 26 2.2.4 Calcium Bromide 28 2.2.5 Calcium Iodide 28 2.2.6 Strontium Chloride 29 2.2.7 Pure Iron 30 2.2.8 Gases 31 2.3 Dry-box. -

Chapter 2 Incandescent Light Bulb
Lamp Contents 1 Lamp (electrical component) 1 1.1 Types ................................................. 1 1.2 Uses other than illumination ...................................... 2 1.3 Lamp circuit symbols ......................................... 2 1.4 See also ................................................ 2 1.5 References ............................................... 2 2 Incandescent light bulb 3 2.1 History ................................................. 3 2.1.1 Early pre-commercial research ................................ 4 2.1.2 Commercialization ...................................... 5 2.2 Tungsten bulbs ............................................. 6 2.3 Efficacy, efficiency, and environmental impact ............................ 8 2.3.1 Cost of lighting ........................................ 9 2.3.2 Measures to ban use ...................................... 9 2.3.3 Efforts to improve efficiency ................................. 9 2.4 Construction .............................................. 10 2.4.1 Gas fill ............................................ 10 2.5 Manufacturing ............................................. 11 2.6 Filament ................................................ 12 2.6.1 Coiled coil filament ...................................... 12 2.6.2 Reducing filament evaporation ................................ 12 2.6.3 Bulb blackening ........................................ 13 2.6.4 Halogen lamps ........................................ 13 2.6.5 Incandescent arc lamps .................................... 14 2.7 Electrical -

AMC Salt Tables
AMC Salt Tables amcmud.com Table of Contents Maximum Densities 2 Potassium Acetate, KC2H3O2 9 Sodium Chloride, NaCl 3 Potassium Formate, KCOOH 10 Potassium Chloride, KC 4 Calcium Bromide, CaBr2 12 Calcium Chloride, CaCl2 5 Sodium Bromide, NaBr 13 Magnesium Chloride, MgCl2 6 Ammonium Chloride, NH4Cl 14 Potassium Sulfate, K2SO4 7 Sodium Nitrate, NaNO3 15 Sodium Formate, NaCOOH 8 Cesium Formate, CsCOOH 16 Maximum Densities 22 1836 2 1670 18 1503 SG 16 1336 lb / gal 14 1169 12 1002 1 835 4 2 3 2 2 2 Cl O 4 3 SO 2 H 2 CsCOOH ride, NH Sodium Chloride, NaCl Sodium Bromide, NaBr Sodium Bromide, Sodium Nitrate, NaNO Calcium Chloride, CaCl Calcium Bromide, CaBr Calcium Bromide, Potassium Chloride, KCl Chloride, KCl Potassium Potassium Sulfate, K Potassium Cesium Formate, Cesium Formate, Sodium Formate, NaCOOH Sodium Formate, Ammonium Chlo Magnesium Chloride, MgCl Potassium Formate, KCOOH Formate, Potassium Potassium Acetate, KC Potassium MAX DENSITY, SG MAX DENSITY, LB / GAL Ammonium Chloride, NH4Cl 1.067 8.90 Potassium Sulfate, K2SO4 1.083 9.03 Potassium Chloride, KCl 1.162 9.69 Sodium Chloride, NaCl 1.197 9.98 Magnesium Chloride, MgCl2 1.276 10.64 Sodium Nitrate, NaNO3 1.320 11.01 Sodium Formate, NaCOOH 1.330 11.06 Potassium Acetate, KC2H3O2 1.393 11.60 Calcium Chloride, CaCl2 1.396 11.64 Sodium Bromide, NaBr 1.530 12.70 Potassium Formate, KCOOH 1.580 13.18 Calcium Bromide, CaBr2 1.830 15.25 Cesium Formate, CsCOOH 2.200 18.36 amcmud.com 2 Sodium Chloride, NaCl WEIGHT DENSITY DENSITY Cl– Na+ NaCl WATER CRYST.