Simultaneous Control of Head and Thoracic Temperature by the Green Darner Dragonfly Anax Junius (Odonata: Aeshnidae)
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Monitoring Dragonfly Migration in North America Protocols for Citizen Scientists
Monitoring Dragonfly Migration in North America Protocols for Citizen Scientists Migratory Dragonfly Partnership Blank on purpose Monitoring Dragonfly Migration in North America Protocols for Citizen Scientists Migratory Dragonfly Partnership Canada • United States • Mexico www.migratorydragonflypartnership.org © 2014 by The Migratory Dragonfly Partnership The Migratory Dragonfly Partnership uses research, citizen science, education, and outreach to under- stand North American dragonfly migration and promote conservation. MDP steering committee members represent a range of organizations, including: Ontario Ministry of Natural Resources; Peggy Notebaert Nature Museum; Pronatura Veracruz; Rutgers University; Slater Museum of Natural History, University of Puget Sound; Smithsonian Conservation Biology Institute; St. Edward's University; U. S. Forest Service International Programs; U. S. Geological Survey; Vermont Center for Ecostudies; and the Xerces Society for Invertebrate Conservation. Migratory Dragonfly Partnership Project Coordinator, Celeste Mazzacano [email protected] 628 NE Broadway, Suite 200, Portland, OR 97232 Tel (855) 232-6639 Fax (503) 233-6794 www.migratorydragonflypartnership.org Acknowledgements Funding for the Migratory Dragonfly Partnership's work is provided by the U.S. Forest Service Inter- national Programs. We thank the photographers who generously allowed use of their images. Copyright of all photographs remains with the photographers. Front and Back Cover Photographs Common Green Darner (Anax junius) male. Photograph © John C. Abbott/Abbott Nature Photography. CONTENTS Summary Page 1 1. Introduction Page 3 1.1 Objectives and Goals Page 3 Box 1: Citizen Science Projects, page 4. 2. Citizen Science Projects Page 5 2.1 Migration Monitoring Page 5 2.1.1 Fall Migration Observations Page 5 - Objectives, page 5. Box 2: MDP Monitoring Projects, page 6. -

Gardening with the Masters Growing, Gardening and Gaining Knowledge August/September 2020
Gardening With The Masters Growing, Gardening and Gaining Knowledge August/September 2020 Editor’s Corner WHAT’S HAPPENING By Marcia Winchester, Cherokee County Master Gardener As the worldwide health concerns continue with the COVID-19 virus, the Cherokee County Master Gardeners also continue to evaluate our monthly events and activities, and continue to follow UGA containers featuring pitcher plant bogs, Extension protocols and state hydrangeas, hostas, and even a number guidelines. of native azaleas. I tuck in a few annuals, and this gives me blooms all year to We continue to ensure the enjoy while sitting at my kitchen table. wellbeing of our members and community. Should we I can view a climbing hydrangea reschedule any of our events, (Schizophragma hydrangeoides) 30 feet you will be notified here, in our up a tulip poplar (Liriodendron tulipifera) newsletter, on our website, or from both the bedroom and family room. on Facebook. I also have a Magnolia liliflora ‘Ann’ that Please feel free to visit any of has beautiful deep pink/purple flowers our web links posted below. that I can see from both rooms. I have They will keep you up to date a grouping of a large clay pot planted with all our activities, events with ferns with a ceramic mushroom and changes, and help you to that I see when I’m practicing yoga. stay informed. A dear friend gave me a combination container with a beautiful 10 foot tall deep pink hibiscus that blooms in mid-summer, and when it stops in late September, an equally tall and dramatic Confederate rose (Hibiscus mutabilis) blooms until frost. -

Odonata: Aeshnidae): a Study of Traits from Larval Development to Adults
EUROPEAN JOURNAL OF ENTOMOLOGYENTOMOLOGY ISSN (online): 1802-8829 Eur. J. Entomol. 116: 269–280, 2019 http://www.eje.cz doi: 10.14411/eje.2019.031 ORIGINAL ARTICLE Biometry of the large dragonfl y Anax imperator (Odonata: Aeshnidae): A study of traits from larval development to adults MARCEAU MINOT 1, MICKAËL LE GALL2 and AURÉLIE HUSTÉ 1 1 Université de Rouen - ECODIV, Bat Blondel, Place Emile Blondel, Mont-Saint-Aignan 76821, France; e-mails: [email protected], [email protected] 2 IRSTEA, UR RIVERLY, Centre de Lyon – Villeurbanne, 69625 Villeurbanne Cedex, France; e-mail: [email protected] Key words. Odonata, Aeshnidae, Anax imperator, body length, body mass, larval rearing, sexual size dimorphism, traits Abstract. Insect larval development affects adult traits but the biometric relationships are usually poorly understood, including large odonates. In this study, measurements of morphological traits of larvae, exuviae and adults of Anax imperator were recorded. They were used to investigate the effects of early development on adult morphology. Results showed an increase in larval length during the fi nal instar and the length of its exuviae signifi cantly exceeded that of the larva. Length and body mass of teneral adults were strongly related to the length of their exuviae. Adult males were signifi cantly longer than adult females, while both had the same body mass at emergence. Length of teneral adults was negatively related to the date of emergence in both sexes. During maturation, body mass of males only increased slightly whereas that of females increased greatly. Mature specimens were also signifi cantly longer than teneral individuals. -

Aquatic and Terrestrial Vegetation Influence
AQUATIC AND TERRESTRIAL VEGETATION INFLUENCE LACUSTRINE DRAGONFLY (ORDER ODONATA) ASSEMBLAGES AT MULTIPLE LIFE STAGES By Alysa J. Remsburg A dissertation submitted in partial fulfillment of the requirements for the degree of Doctor of Philosophy (Zoology) at the UNIVERSITY OF WISCONSIN – MADISON 2007 i ACKNOWLEDGMENTS Reflecting on the contributions of my colleagues and friends during my graduate studies gives me a strong sense of gratitude for the community of support that I have enjoyed. The people who surround and support me deserve more thanks than I can describe here. Friends and family have supported my graduate studies by generously accommodating my tight schedule and warmly offering encouragement throughout the process. Monica Turner guided my graduate studies in numerous ways. It was her trust in my abilities and willingness to learn about a new study organism that first made this research possible. She encouraged me to pursue the research questions that most interested and inspired me, although it meant charting territory that was new to both of us. Monica served as the ideal mentor for me by requiring clear communication, modeling an efficient and balanced work ethic, providing critical reviews, and listening compassionately. This research benefited from the expertise and generosity of outstanding Wisconsin ecologists. Members of my graduate research committee, Steve Carpenter, Claudio Gratton, Tony Ives, Bobbi Peckarsky, and Joy Zedler, all offered useful suggestions and critiques on experimental design, pressing research questions, and the manuscripts. Cecile Ane provided additional statistical advice and smiles. Bill Smith, Bob DuBois, and Robert Bohanan answered (or reassured me that I should try to answer) many questions about field methods, Odonata biology, and species identification. -

Green Darner Anax Junius
TEXAS PARKS AND WILDLIFE Common Green Darner Anax junius TPWD PHOTO Dragonflies haven’t changed much over the last 300 million years. Today they are one of the most easily recognized of all insects. The common green darner is the largest, most abundant and widespread dragonfly species in North America. Common Green Darner Anax junius APPEARANCE HABITAT Length: 3 inches (7.6cm) Common green darners prefer permanent and temporary ponds, Wingspan: 4.5 inches (11.4cm) lakes, bays, estuaries and slow-moving streams and riparian areas (land adjacent to a body of water). Distinguishing Characteristics • Green head and thorax • Abdomen is yellow and brown on females; long, slender and BEHAVIOR bluish on males • Large compound eyes During the reproductive stage, the common green darner seeks • Strong jaws fresh water ponds. Males return first, then females who have • Spiny legs already developed a batch of eggs. If the female is receptive, • Wings clear with yellowish tint toward tips and strongly mating begins. They often mate in flight. Immediately after mat- veined with net-like pattern ing, common green darner females lay their eggs, one at a time, in tiny slits in submerged aquatic plants. Males work hard to protect their territories. Some have been clocked chasing intrud- LIFE HISTORY ers away at 35 miles per hour. Common green darners are one of the few dragonflies that migrate in the spring and fall. Range: North America, the West Indies and Scientists believe that they migrate with seasonal warm fronts. South America Diet: Larvae eat fish eggs, tadpoles and other small aquatic animals. -

Creating Economically and Ecologically Sustainable Pollinator Habitat District 2 Demonstration Research Project Summary Updated for Site Visit in April 2019
Creating Economically and Ecologically Sustainable Pollinator Habitat District 2 Demonstration Research Project Summary Updated for Site Visit in April 2019 The PIs are most appreciative for identification assistance provided by: Arian Farid and Alan R. Franck, Director and former Director, resp., University of South Florida Herbarium, Tampa, FL; Edwin Bridges, Botanical and Ecological Consultant; Floyd Griffith, Botanist; and Eugene Wofford, Director, University of Tennessee Herbarium, Knoxville, TN Investigators Rick Johnstone and Robin Haggie (IVM Partners, 501-C-3 non-profit; http://www.ivmpartners.org/); Larry Porter and John Nettles (ret.), District 2 Wildflower Coordinator; Jeff Norcini, FDOT State Wildflower Specialist Cooperator Rick Owen (Imperiled Butterflies of Florida Work Group – North) Objective Evaluate a cost-effective strategy for creating habitat for pollinators/beneficial insects in the ROW beyond the back-slope. Rationale • Will aid FDOT in developing a strategy to create pollinator habitat per the federal BEE Act and FDOT’s Wildflower Program • Will demonstrate that FDOT can simultaneously • Create sustainable pollinator habitat in an economical and ecological manner • Reduce mowing costs • Part of national effort coordinated by IVM Partners, who has • Established or will establish similar projects on roadside or utility ROWS in Alabama, Arkansas, Maryland, New Mexico, Oklahoma, Idaho, Montana, Virginia, West Virginia, and Tennessee; studies previously conducted in Arizona, Delaware, Michigan, and New Jersey • Developed partnerships with US Fish & Wildlife Service, Army Corps of Engineers, US Geological Survey, New Jersey Institute of Technology, Rutgers University, Chesapeake Bay Foundation, Chesapeake Wildlife Heritage, The Navajo Nation, The Wildlife Habitat Council, The Pollinator Partnership, Progressive Solutions, Bayer Crop Sciences, Universities of Maryland, Ohio, West Virginia, and the EPA. -

Barcode of Life Data Systems (BOLD) Versus Genbank Molecular Identification of a Dragonfly from the UAE in Comparison to the Morphological Identification
OnLine Journal of Biological Sciences Original Research Paper Barcode of Life Data Systems (BOLD) Versus GenBank Molecular Identification of a Dragonfly from the UAE in Comparison to the Morphological Identification 1Noora Almansoori, 1,2Mohamed Rizk Enan and 1Mohammad Ali Al-Deeb 1Department of Biology, United Arab Emirates University, Al-Ain, UAE 2Agricultural Research Center, Agricultural Genetic Engineering Research Institute, Giza, Egypt Article history Abstract: Dragonflies are insects in the order Odonata. They inhabit Received: 26-09-2019 freshwater ecosystems and are found in the UAE. To date, few checklists Revised: 19-11-2019 have been published for the local dragonflies and the used identification Accepted: 29-11-2019 keys are not comprehensive of Arabia. The aim of this study was to provide a molecular identification of a dragonfly based on the mitochondrial Corresponding Author: Cytochrome c Oxidase subunit I (COI) gene using the National Center for Mohammad Ali Al-Deeb, Biotechnology Information (NCBI) database and the Barcode of Life Data Department of Biology, United Systems (BOLD) in comparison with the morphology. The insect’s DNA Arab Emirates University, Al- was extracted and the PCR was performed on the target gene. The insect Ain, UAE Email: [email protected] was identified initially as Anax imperator based on the NCBI database and as Anax parthenope based on the BOLD. However, the morphological identification was in agreement with the one produced by the BOLD. The results of this study is a demonstration of how, in some cases, the DNA- based identification does not provide a conclusive species designation and that a morphology-based identification is needed. -

Identification Guide to the Australian Odonata Australian the to Guide Identification
Identification Guide to theAustralian Odonata www.environment.nsw.gov.au Identification Guide to the Australian Odonata Department of Environment, Climate Change and Water NSW Identification Guide to the Australian Odonata Department of Environment, Climate Change and Water NSW National Library of Australia Cataloguing-in-Publication data Theischinger, G. (Gunther), 1940– Identification Guide to the Australian Odonata 1. Odonata – Australia. 2. Odonata – Australia – Identification. I. Endersby I. (Ian), 1941- . II. Department of Environment and Climate Change NSW © 2009 Department of Environment, Climate Change and Water NSW Front cover: Petalura gigantea, male (photo R. Tuft) Prepared by: Gunther Theischinger, Waters and Catchments Science, Department of Environment, Climate Change and Water NSW and Ian Endersby, 56 Looker Road, Montmorency, Victoria 3094 Published by: Department of Environment, Climate Change and Water NSW 59–61 Goulburn Street Sydney PO Box A290 Sydney South 1232 Phone: (02) 9995 5000 (switchboard) Phone: 131555 (information & publication requests) Fax: (02) 9995 5999 Email: [email protected] Website: www.environment.nsw.gov.au The Department of Environment, Climate Change and Water NSW is pleased to allow this material to be reproduced in whole or in part, provided the meaning is unchanged and its source, publisher and authorship are acknowledged. ISBN 978 1 74232 475 3 DECCW 2009/730 December 2009 Printed using environmentally sustainable paper. Contents About this guide iv 1 Introduction 1 2 Systematics -

© 2016 David Paul Moskowitz ALL RIGHTS RESERVED
© 2016 David Paul Moskowitz ALL RIGHTS RESERVED THE LIFE HISTORY, BEHAVIOR AND CONSERVATION OF THE TIGER SPIKETAIL DRAGONFLY (CORDULEGASTER ERRONEA HAGEN) IN NEW JERSEY By DAVID P. MOSKOWITZ A dissertation submitted to the Graduate School-New Brunswick Rutgers, The State University of New Jersey In partial fulfillment of the requirements For the degree of Doctor of Philosophy Graduate Program in Entomology Written under the direction of Dr. Michael L. May And approved by _____________________________________ _____________________________________ _____________________________________ _____________________________________ New Brunswick, New Jersey January, 2016 ABSTRACT OF THE DISSERTATION THE LIFE HISTORY, BEHAVIOR AND CONSERVATION OF THE TIGER SPIKETAIL DRAGONFLY (CORDULEGASTER ERRONEA HAGEN) IN NEW JERSEY by DAVID PAUL MOSKOWITZ Dissertation Director: Dr. Michael L. May This dissertation explores the life history and behavior of the Tiger Spiketail dragonfly (Cordulegaster erronea Hagen) and provides recommendations for the conservation of the species. Like most species in the genus Cordulegaster and the family Cordulegastridae, the Tiger Spiketail is geographically restricted, patchily distributed with its range, and a habitat specialist in habitats susceptible to disturbance. Most Cordulegastridae species are also of conservation concern and the Tiger Spiketail is no exception. However, many aspects of the life history of the Tiger Spiketail and many other Cordulegastridae are poorly understood, complicating conservation strategies. In this dissertation, I report the results of my research on the Tiger Spiketail in New Jersey. The research to investigate life history and behavior included: larval and exuvial sampling; radio- telemetry studies; marking-resighting studies; habitat analyses; observations of ovipositing females and patrolling males, and the presentation of models and insects to patrolling males. -

Cumulative Index of ARGIA and Bulletin of American Odonatology
Cumulative Index of ARGIA and Bulletin of American Odonatology Compiled by Jim Johnson PDF available at http://odonata.bogfoot.net/docs/Argia-BAO_Cumulative_Index.pdf Last updated: 14 February 2021 Below are titles from all issues of ARGIA and Bulletin of American Odonatology (BAO) published to date by the Dragonfly Society of the Americas. The purpose of this listing is to facilitate the searching of authors and title keywords across all issues in both journals, and to make browsing of the titles more convenient. PDFs of ARGIA and BAO can be downloaded from https://www.dragonflysocietyamericas.org/en/publications. The most recent three years of issues for both publications are only available to current members of the Dragonfly Society of the Americas. Contact Jim Johnson at [email protected] if you find any errors. ARGIA 1 (1–4), 1989 Welcome to the Dragonfly Society of America Cook, C. 1 Society's Name Revised Cook, C. 2 DSA Receives Grant from SIO Cook, C. 2 North and Central American Catalogue of Odonata—A Proposal Donnelly, T.W. 3 US Endangered Species—A Request for Information Donnelly, T.W. 4 Odonate Collecting in the Peruvian Amazon Dunkle, S.W. 5 Collecting in Costa Rica Dunkle, S.W. 6 Research in Progress Garrison, R.W. 8 Season Summary Project Cook, C. 9 Membership List 10 Survey of Ohio Odonata Planned Glotzhober, R.C. 11 Book Review: The Dragonflies of Europe Cook, C. 12 Book Review: Dragonflies of the Florida Peninsula, Bermuda and the Bahamas Cook, C. 12 Constitution of the Dragonfly Society of America 13 Exchanges and Notices 15 General Information About the Dragonfly Society of America (DSA) Cook, C. -

Observations on Egg Parasitism of Aeshna Tuberculifera (Odonata: Aeshnidae) by Eulophidae, Trichogrammatidae and Mymaridae (Hymenoptera) in Alger County, Michigan
The Great Lakes Entomologist Volume 46 Numbers 3 & 4 - Fall/Winter 2013 Numbers 3 & Article 1 4 - Fall/Winter 2013 October 2013 Observations on Egg Parasitism of Aeshna Tuberculifera (Odonata: Aeshnidae) by Eulophidae, Trichogrammatidae and Mymaridae (Hymenoptera) in Alger County, Michigan Burton C. Cebulski Mark F. O'Brien University of Michigan Follow this and additional works at: https://scholar.valpo.edu/tgle Part of the Entomology Commons Recommended Citation Cebulski, Burton C. and O'Brien, Mark F. 2013. "Observations on Egg Parasitism of Aeshna Tuberculifera (Odonata: Aeshnidae) by Eulophidae, Trichogrammatidae and Mymaridae (Hymenoptera) in Alger County, Michigan," The Great Lakes Entomologist, vol 46 (2) Available at: https://scholar.valpo.edu/tgle/vol46/iss2/1 This Peer-Review Article is brought to you for free and open access by the Department of Biology at ValpoScholar. It has been accepted for inclusion in The Great Lakes Entomologist by an authorized administrator of ValpoScholar. For more information, please contact a ValpoScholar staff member at [email protected]. Cebulski and O'Brien: Observations on Egg Parasitism of <i>Aeshna Tuberculifera</i> (Od 2013 THE GREAT LAKES ENTOMOLOGIST 145 Observations on Egg Parasitism of Aeshna tuberculifera (Odonata: Aeshnidae) by Eulophidae, Trichogrammatidae and Mymaridae (Hymenoptera) in Alger County, Michigan Burton C. Cebulski1 and Mark F. O'Brien2 Abstract Egg parasitoids were reared from a population of Aeshna tuberculifera (Odonata: Aeshnidae) in the Kingston Lake area of Alger County, Michigan, from 1983-2005. Leaves of Iris versicolor were repeatedly used for oviposition during the period of observation, with the result that just a few leaves were targeted by different females. -
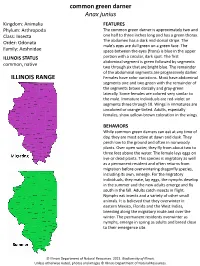
Common Green Darner Anax Junius ILLINOIS RANGE
common green darner Anax junius Kingdom: Animalia FEATURES Phylum: Arthropoda The common green darner is approximately two and Class: Insecta one half to three inches long and has a green thorax. Order: Odonata The abdomen has a dark mid-dorsal stripe. The male’s eyes are dull green on a green face. The Family: Aeshnidae space between the eyes (frons) is blue in the upper ILLINOIS STATUS portion with a circular, dark spot. The first abdominal segment is green followed by segments common, native two through six that are bright blue. The remainder of the abdominal segments are progressively darker. ILLINOIS RANGE Females have color variations. Most have abdominal segments one and two green with the remainder of the segments brown dorsally and gray-green laterally. Some females are colored very similar to the male. Immature individuals are red-violet on segments three through 10. Wings in immatures are uncolored or orange-tinted. Adults, especially females, show yellow-brown coloration in the wings. BEHAVIORS While common green darners can eat at any time of day, they are most active at dawn and dusk. They perch low to the ground and often in nonwoody plants. Over open water, they fly from about two to three feet above the water. The female lays eggs on live or dead plants. This species is migratory as well as a permanent resident and often returns from migration before overwintering dragonfly species, including its own, emerge. For the migratory individuals, they mate, lay eggs, the nymphs develop in the summer and the new adults emerge and fly south in the fall.